

Multiple-myeloma-related WHSC1/MMSET isoform RE-IIBP is a histone methyltransferase with transcriptional repression activity
- PMID: 18172012
- PMCID: PMC2268398
- DOI: 10.1128/MCB.02130-07
Multiple-myeloma-related WHSC1/MMSET isoform RE-IIBP is a histone methyltransferase with transcriptional repression activity
Abstract
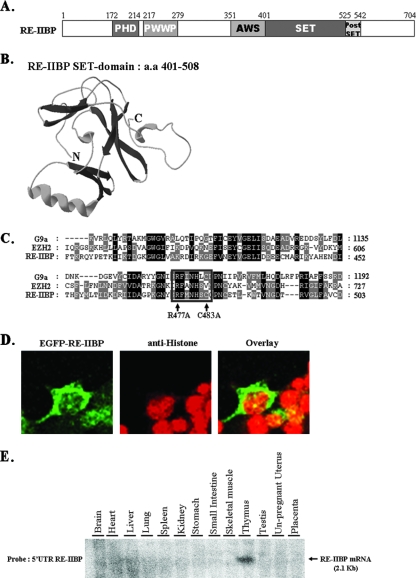
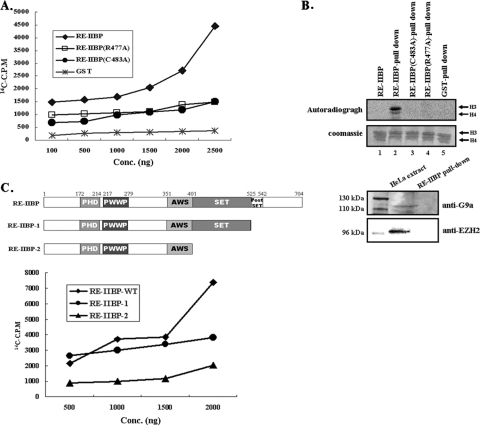
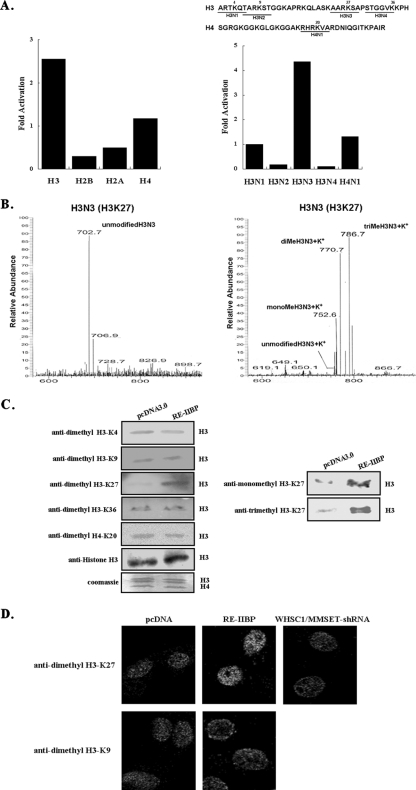
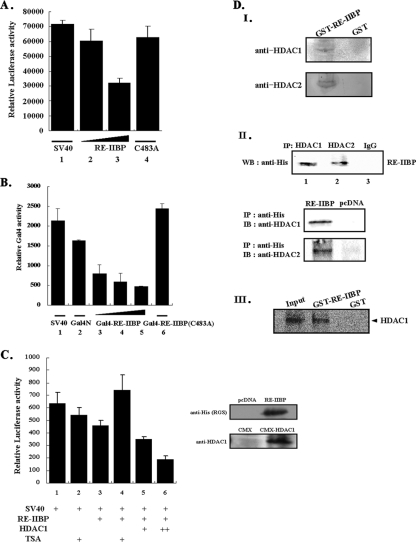
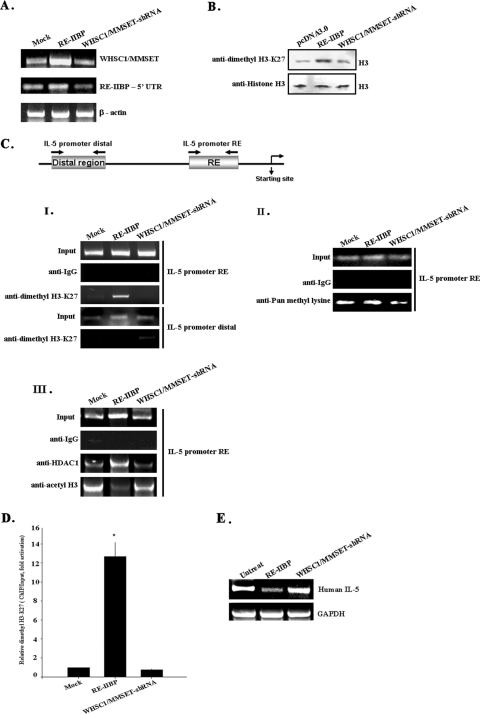
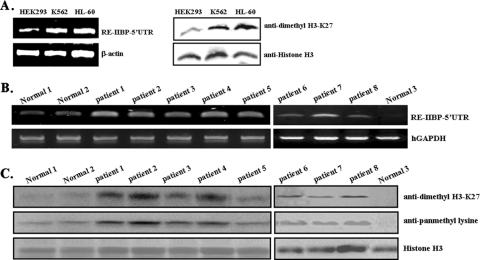
Histone methylation is crucial for transcriptional regulation and chromatin remodeling. It has been suggested that the SET domain containing protein RE-IIBP (interleukin-5 [IL-5] response element II binding protein) may perform a function in the carcinogenesis of certain tumor types, including myeloma. However, the pathogenic role of RE-IIBP in these diseases remains to be clearly elucidated. In this study, we have conducted an investigation into the relationship between the histone-methylating activity of RE-IIBP and transcriptional regulation. Here, we report that RE-IIBP is up-regulated in the blood cells of leukemia patients, and we characterized the histone H3 lysine 27 (H3-K27) methyltransferase activity of RE-IIBP. Point mutant analysis revealed that SET domain cysteine 483 and arginine 477 are critical residues for the histone methyltransferase (HMTase) activity of RE-IIBP. RE-IIBP also represses basal transcription via histone deacetylase (HDAC) recruitment, which may be mediated by H3-K27 methylation. In the chromatin immunoprecipitation assays, we showed that RE-IIBP overexpression induces histone H3-K27 methylation, HDAC recruitment, and histone H3 hypoacetylation on the IL-5 promoter and represses expression. Conversely, short hairpin RNA-mediated knockdown of RE-IIBP reduces histone H3-K27 methylation and HDAC occupancy around the IL-5 promoter. These data illustrate the important regulatory role of RE-IIBP in transcriptional regulation, thereby pointing to the important role of HMTase activity in carcinogenesis.
Figures
References
-
- Ayton, P. M., and M. L. Cleary. 2001. Molecular mechanisms of leukemogenesis mediated by MLL fusion proteins. Oncogene 205695-5707. - PubMed
-
- Cao, R., L. Wang, H. Wang, L. Xia, H. Erdjument-Bromage, P. Tempst, R. S. Jones, and Y. Zhang. 2002. Role of histone H3 lysine 27 methylation in Polycomb-group silencing. Science 2981039-1043. - PubMed
-
- Daser, A., and T. H. Rabbitts. 2004. Extending the repertoire of the mixed-lineage leukemia gene MLL in leukemogenesis. Genes Dev. 18965-974. - PubMed
-
- Fischle, W., Y. Wang, and C. D. Allis. 2003. Binary switches and modification cassettes in histone biology and beyond. Nature 425475-479. - PubMed
-
- Garlisi, C. G., A. S. Uss, H. Xiao, F. Tian, K. E. Sheridan, L. Wang, M. Motasim Billah, R. W. Egan, K. S. Stranick, and S. P. Umland. 2001. A unique mRNA initiated within a middle intron of WHSC1/MMSET encodes a DNA binding protein that suppresses human IL-5 transcription. Am. J. Respir. Cell Mol. Biol. 2490-98. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases