

Review
doi: 10.1101/gad.1634508.
New ways to meet your (3') end oligouridylation as a step on the path to destruction
Affiliations
- PMID: 18172159
- PMCID: PMC2731568
- DOI: 10.1101/gad.1634508
Item in Clipboard
Review
New ways to meet your (3') end oligouridylation as a step on the path to destruction
Genes Dev.
.
No abstract available
Figures
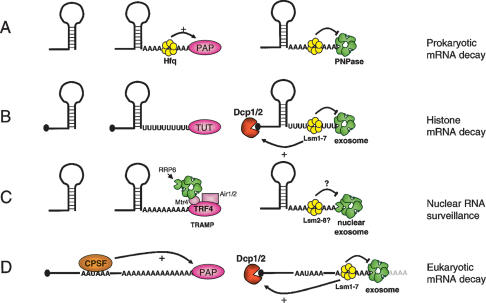
3′ Single-stranded extensions lead to similar degradation pathways in prokaryotes and eukaryotes. (A) Prokaryotic mRNAs degrade from the 3′ end through polyadenylation by PAP1 and subsequent 3′–5′ exonucleolytic decay mediated either by PNPase as shown here, or other 3′–5′ exonucleases. Hfq, an Sm-like protein, associates with the 3′ poly(A) tract and modulates PAP and PNPase activity. (B) Histone mRNAs undergo oligouridylation by a cytoplasmic terminal uridyl transferase at the end of S phase. This leads to association of Lsm11–7, and recruitment of the decapping and 5′–3′ decay machinery. Decay also occurs 3′–5′ by the exosome. How Lsm1–7 association influences exosome activity is unclear, although there is evidence for an inhibitory role (see the text). (C) In yeast, aberrant nuclear RNAs undergo polyadenylation by the TRAMP complex, which contains the Trf4 noncanonical PAP as well as a helicase (Mtr4) and an RNA-binding protein (Air1 or Air2). TRAMP recruits the nuclear exosome (containing Rrp6) to the polyadenylated RNA leading to rapid degradation. It is not clear whether the nuclear Lsm complex (Lsm2–8) influences this process. (D) Eukaryotic mRNAs undergo polyadenylation in the nucleus in a cotranscriptional process involving the CPSF and nuclear PAP, among other factors. In the cytoplasm, these mRNAs undergo degradation that is initiated by removal of the majority of the poly(A) tail (not shown). Lsm1–7 is thought to associate with the remaining 3′ oligo(A) tract and recruit the decapping machinery to induce 5′–3′ decay. In addition, the exosome degrades the transcript 3′–5′. The interplay between the exosome and Lsm1–7 is not clear, although evidence suggests that Lsm inhibits exosome activity.
Comment on
-
Degradation of histone mRNA requires oligouridylation followed by decapping and simultaneous degradation of the mRNA both 5' to 3' and 3' to 5'.Genes Dev. 2008 Jan 1;22(1):50-65. doi: 10.1101/gad.1622708. Genes Dev. 2008. PMID: 18172165 Free PMC article.
References
-
- Barnard D.C., Ryan K., Manley J.L., Richter J.D. Symplekin and xGLD-2 are required for CPEB-mediated cytoplasmic polyadenylation. Cell. 2004;119:641–651. - PubMed
-
- Buhler M., Haas W., Gygi S.P., Moazed D. RNAi-dependent and -independent RNA turnover mechanisms contribute to heterochromatic gene silencing. Cell. 2007;129:707–721. - PubMed
-
- Carpousis A.J. The RNA degradosome of Escherichia coli: An mRNA-degrading machine assembled on RNase E. Annu. Rev. Microbiol. 2007;61:71–87. - PubMed
-
- Chen C.C., Simard M.J., Tabara H., Brownell D.R., McCollough J.A., Mello C.C. A member of the polymerase β nucleotidyltransferase superfamily is required for RNA interference in C. elegans. Curr. Biol. 2005;15:378–383. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources