

APC/C-Cdh1-mediated degradation of the Polo kinase Cdc5 promotes the return of Cdc14 into the nucleolus
- PMID: 18172166
- PMCID: PMC2151016
- DOI: 10.1101/gad.1601308
APC/C-Cdh1-mediated degradation of the Polo kinase Cdc5 promotes the return of Cdc14 into the nucleolus
Abstract
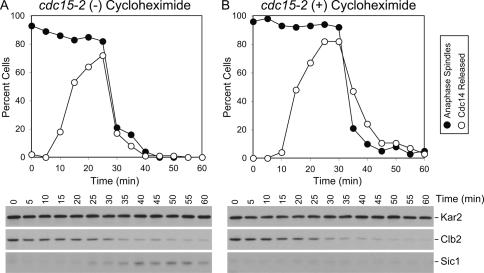
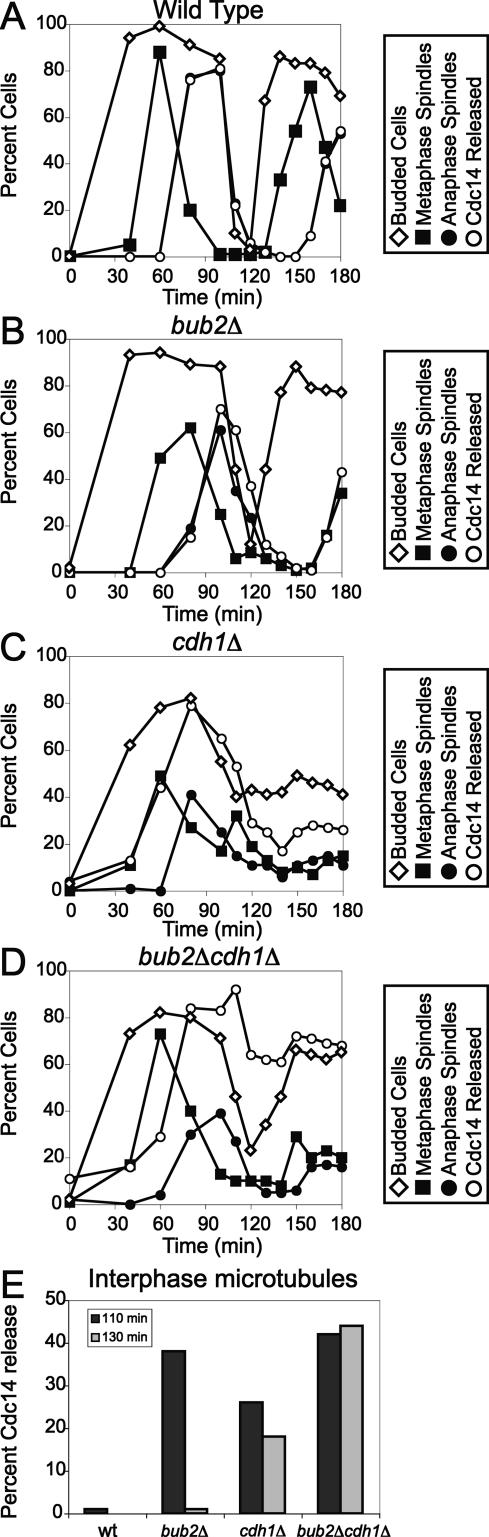
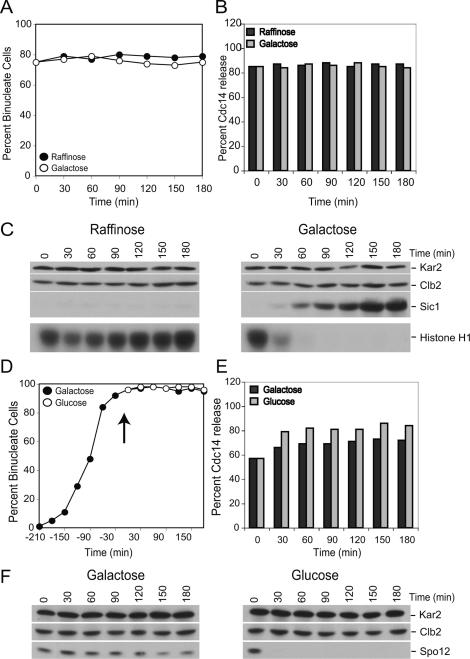
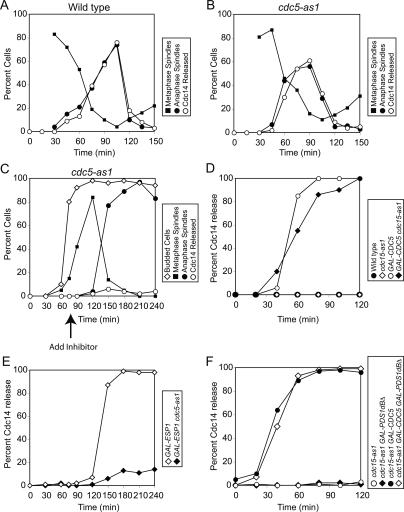
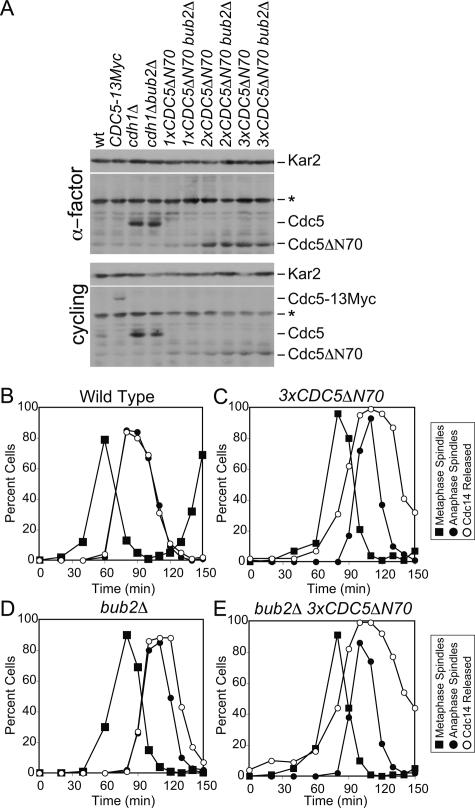
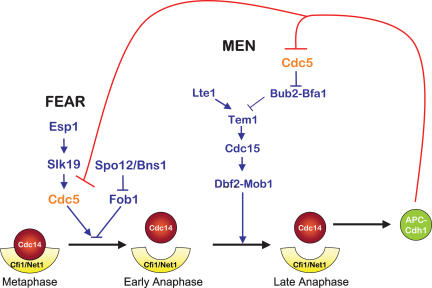
In the budding yeast Saccharomyces cerevisiae, the protein phosphatase Cdc14 triggers exit from mitosis by promoting the inactivation of cyclin-dependent kinases (CDKs). Cdc14's activity is controlled by Cfi1/Net1, which holds and inhibits the phosphatase in the nucleolus from G1 until metaphase. During anaphase, two regulatory networks, the Cdc14 Early Anaphase Release (FEAR) network and the Mitotic Exit Network (MEN), promote the dissociation of Cdc14 from its inhibitor, allowing the phosphatase to reach its targets throughout the cell. The molecular circuits that trigger the return of Cdc14 into the nucleolus after the completion of exit from mitosis are not known. Here we show that activation of a ubiquitin ligase known as the Anaphase-Promoting Complex or Cyclosome (APC/C) bound to the specificity factor Cdh1 triggers the degradation of the Polo kinase Cdc5, a key factor in releasing Cdc14 from its inhibitor in the nucleolus.
Figures

References
-
- Amon A., Irniger S., Nasmyth K. Closing the cell cycle circle in yeast: G2 cyclin proteolysis initiated at mitosis persists until the activation of G1 cyclins in the next cycle. Cell. 1994;77:1037–1050. - PubMed
-
- Azzam R., Chen S.L., Shou W., Mah A.S., Alexandru G., Nasmyth K., Annan R.S., Carr S.A., Deshaies R.J. Phosphorylation by cyclin B–Cdk underlies release of mitotic exit activator Cdc14 from the nucleolus. Science. 2004;305:516–519. - PubMed
-
- Bachmair A., Finley D., Varshavsky A. In vivo half-life of a protein is a function of its amino-terminal residue. Science. 1986;234:179–186. - PubMed
-
- Bardin A.J., Amon A. Men and sin: What’s the difference? Nat. Rev. Mol. Cell Biol. 2001;2:815–826. - PubMed
-
- Bishop A.C., Buzko O., Shokat K.M. Magic bullets for protein kinases. Trends Cell Biol. 2001;11:167–172. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous