

A chemokine-degrading extracellular protease made by group A Streptococcus alters pathogenesis by enhancing evasion of the innate immune response
- PMID: 18174342
- PMCID: PMC2258835
- DOI: 10.1128/IAI.01354-07
A chemokine-degrading extracellular protease made by group A Streptococcus alters pathogenesis by enhancing evasion of the innate immune response
Abstract
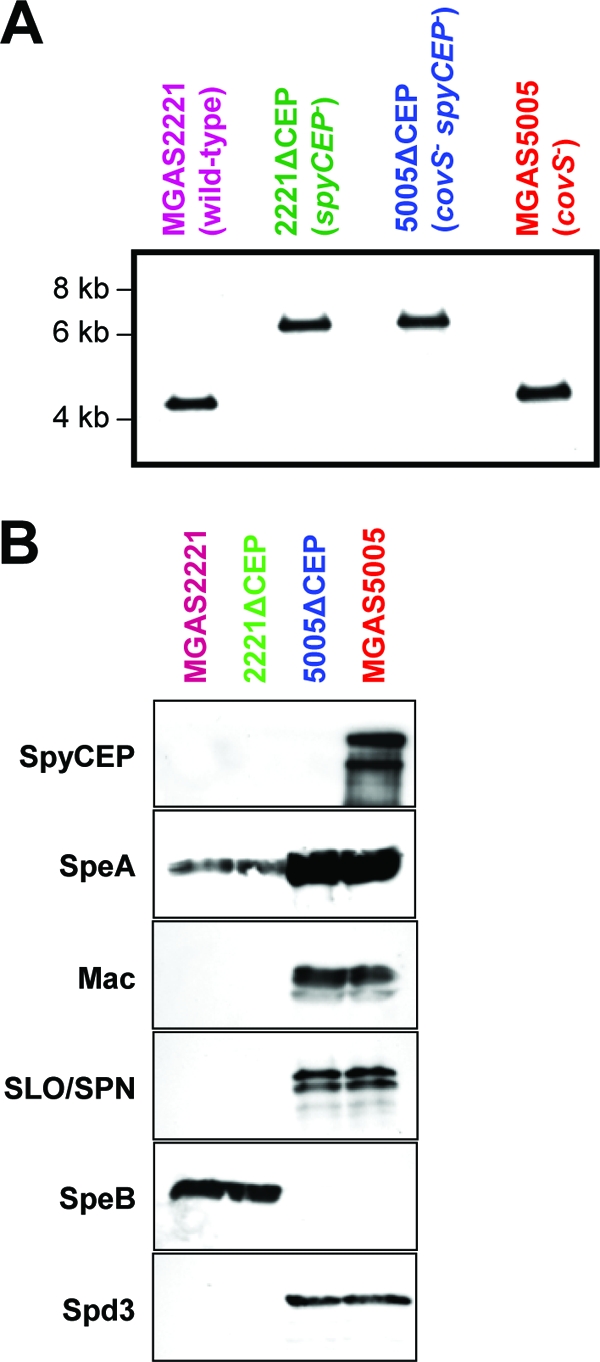
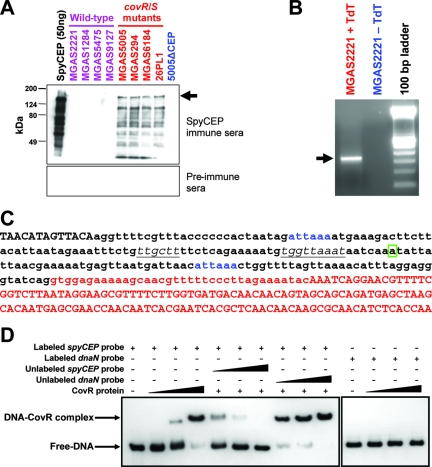
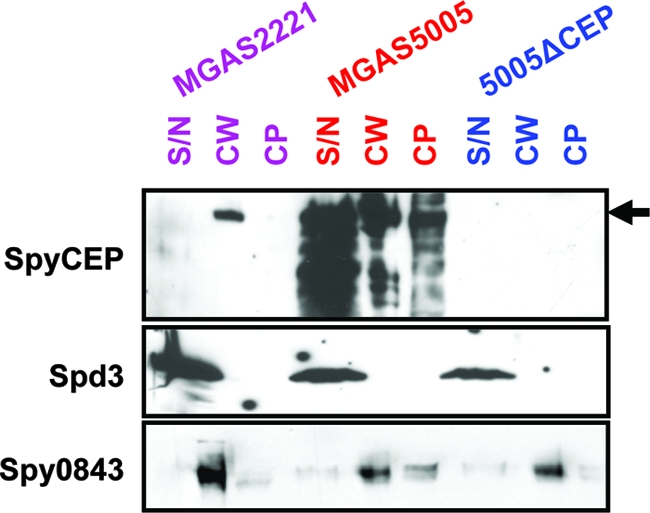
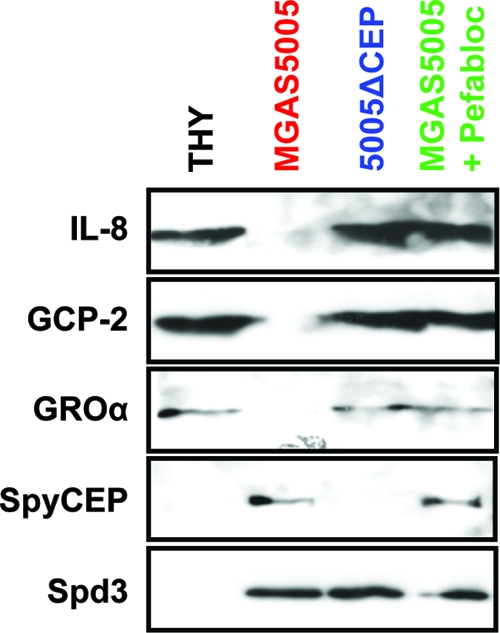
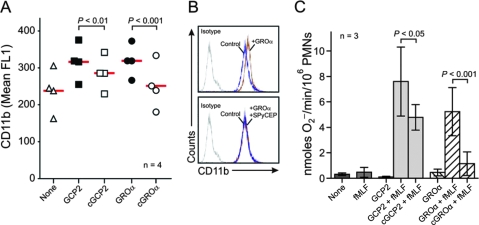
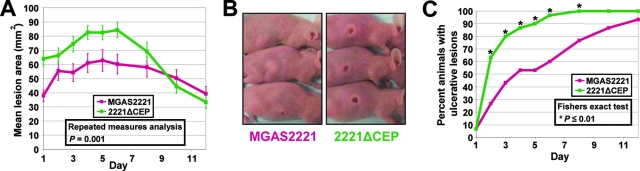
Circumvention of the host innate immune response is critical for bacterial pathogens to infect and cause disease. Here we demonstrate that the group A Streptococcus (GAS; Streptococcus pyogenes) protease SpyCEP (S. pyogenes cell envelope protease) cleaves granulocyte chemotactic protein 2 (GCP-2) and growth-related oncogene alpha (GROalpha), two potent chemokines made abundantly in human tonsils. Cleavage of GCP-2 and GROalpha by SpyCEP abrogated their abilities to prime neutrophils for activation, detrimentally altering the innate immune response. SpyCEP expression is negatively regulated by the signal transduction system CovR/S. Purified recombinant CovR bound the spyCEP gene promoter region in vitro, indicating direct regulation. Immunoreactive SpyCEP protein was present in the culture supernatants of covR/S mutant GAS strains but not in supernatants from wild-type strains. However, wild-type GAS strains do express SpyCEP, where it is localized to the cell wall. Strain MGAS2221, an organism representative of the highly virulent and globally disseminated M1T1 GAS clone, differed significantly from its isogenic spyCEP mutant derivative strain in a mouse soft tissue infection model. Interestingly, and in contrast to previous studies, the isogenic mutant strain generated lesions of larger size than those formed following infection with the parent strain. The data indicate that SpyCEP contributes to GAS virulence in a strain- and disease-dependent manner.
Figures
References
-
- Brinkmann, V., U. Reichard, C. Goosmann, B. Fauler, Y. Uhlemann, D. S. Weiss, Y. Weinrauch, and A. Zychlinsky. 2004. Neutrophil extracellular traps kill bacteria. Science 3031532-1535. - PubMed
-
- Carlsson, F., C. Sandin, and G. Lindahl. 2005. Human fibrinogen bound to Streptococcus pyogenes M protein inhibits complement deposition via the classical pathway. Mol. Microbiol. 5628-39. - PubMed
-
- Cockerill, F. R., III, R. L. Thompson, J. M. Musser, P. M. Schlievert, J. Talbot, K. E. Holley, W. S. Harmsen, D. M. Ilstrup, P. C. Kohner, M. H. Kim, B. Frankfort, J. M. Manahan, J. M. Steckelberg, F. Roberson, W. R. Wilson, et al. 1998. Molecular, serological, and clinical features of 16 consecutive cases of invasive streptococcal disease. Clin. Infect. Dis. 261448-1458. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
