

Determinants of substrate specificity in RNA-dependent nucleotidyl transferases
- PMID: 18177750
- PMCID: PMC2676681
- DOI: 10.1016/j.bbagrm.2007.12.003
Determinants of substrate specificity in RNA-dependent nucleotidyl transferases
Abstract
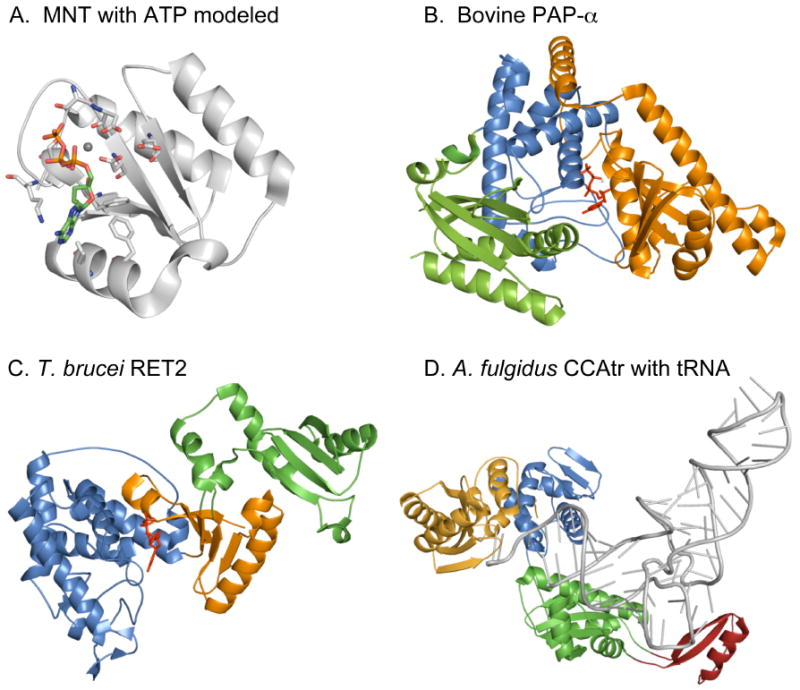
Poly(A) polymerases were identified almost 50 years ago as enzymes that add multiple AMP residues to the 3' ends of primer RNAs without use of a template from ATP as cosubstrate and with release of pyrophosphate. Based on sequence homology of a signature motif in the catalytic domain, poly(A) polymerases were later found to belong to a superfamily of nucleotidyl transferases acting on a very diverse array of substrates. Enzymes belonging to the superfamily can add from single nucleotides of AMP, CMP or UMP to RNA, antibiotics and proteins but also homopolymers of many hundred residues to the 3' ends of RNA molecules. The recently reported structures of several nucleotidyl transferases facilitate the study of the catalytic mechanisms of these very diverse enzymes. Numerous structures of CCA-adding enzymes have now revealed all steps in the formation of a CCA tail at the 3' end of tRNAs. In addition, structures of poly(A) polymerases and uridylyl transferases are now available as binary and ternary complexes with incoming nucleotide and RNA primer. Some of these proteins undergo significant conformational changes after substrate binding. This is proposed to be an indication for an induced fit mechanism that drives substrate selection and leads to catalysis. Insights from recent structures of ternary complexes indicate an important role for the primer molecule in selecting the incoming nucleotide.
Figures




Similar articles
-
RNA-specific ribonucleotidyl transferases.RNA. 2007 Nov;13(11):1834-49. doi: 10.1261/rna.652807. Epub 2007 Sep 13. RNA. 2007. PMID: 17872511 Free PMC article. Review.
-
Dual role of the RNA substrate in selectivity and catalysis by terminal uridylyl transferases.Proc Natl Acad Sci U S A. 2007 Sep 11;104(37):14634-9. doi: 10.1073/pnas.0704259104. Epub 2007 Sep 4. Proc Natl Acad Sci U S A. 2007. PMID: 17785418 Free PMC article.
-
Metal-ion-dependent catalysis and specificity of CCA-adding enzymes: a comparison of two classes.Biochemistry. 2005 Sep 27;44(38):12849-59. doi: 10.1021/bi0509402. Biochemistry. 2005. PMID: 16171400
-
Domain movements during CCA-addition: a new function for motif C in the catalytic core of the human tRNA nucleotidyltransferases.RNA Biol. 2015;12(4):435-46. doi: 10.1080/15476286.2015.1018502. RNA Biol. 2015. PMID: 25849199 Free PMC article.
-
tRNA-nucleotidyltransferases: highly unusual RNA polymerases with vital functions.FEBS Lett. 2010 Jan 21;584(2):297-302. doi: 10.1016/j.febslet.2009.10.078. FEBS Lett. 2010. PMID: 19883645 Review.
Cited by
-
The novel poly(A) polymerase Star-PAP is a signal-regulated switch at the 3'-end of mRNAs.Adv Biol Regul. 2013 Jan;53(1):64-76. doi: 10.1016/j.jbior.2012.10.004. Epub 2012 Oct 13. Adv Biol Regul. 2013. PMID: 23306079 Free PMC article. Review.
-
The role of sequence context, nucleotide pool balance and stress in 2'-deoxynucleotide misincorporation in viral, bacterial and mammalian RNA.Nucleic Acids Res. 2016 Oct 14;44(18):8962-8975. doi: 10.1093/nar/gkw572. Epub 2016 Jun 30. Nucleic Acids Res. 2016. PMID: 27365049 Free PMC article.
-
Structure and function of the polymerase core of TRAMP, a RNA surveillance complex.Proc Natl Acad Sci U S A. 2010 Aug 24;107(34):15045-50. doi: 10.1073/pnas.1003505107. Epub 2010 Aug 9. Proc Natl Acad Sci U S A. 2010. PMID: 20696927 Free PMC article.
-
Protein-primed DNA homopolymer synthesis by an antiviral reverse transcriptase.bioRxiv [Preprint]. 2025 Mar 25:2025.03.24.645077. doi: 10.1101/2025.03.24.645077. bioRxiv. 2025. Update in: Nature. 2025 Jul;643(8074):1352-1362. doi: 10.1038/s41586-025-09179-5. PMID: 40196691 Free PMC article. Updated. Preprint.
-
TRAMP assembly alters the conformation and RNA binding of Mtr4 and Trf4-Air2.Proc Natl Acad Sci U S A. 2025 Jan 7;122(1):e2414980121. doi: 10.1073/pnas.2414980121. Epub 2025 Jan 3. Proc Natl Acad Sci U S A. 2025. PMID: 39752526 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
