

Krp1 (Sarcosin) promotes lateral fusion of myofibril assembly intermediates in cultured mouse cardiomyocytes
- PMID: 18178185
- PMCID: PMC2275804
- DOI: 10.1016/j.yexcr.2007.12.009
Krp1 (Sarcosin) promotes lateral fusion of myofibril assembly intermediates in cultured mouse cardiomyocytes
Abstract
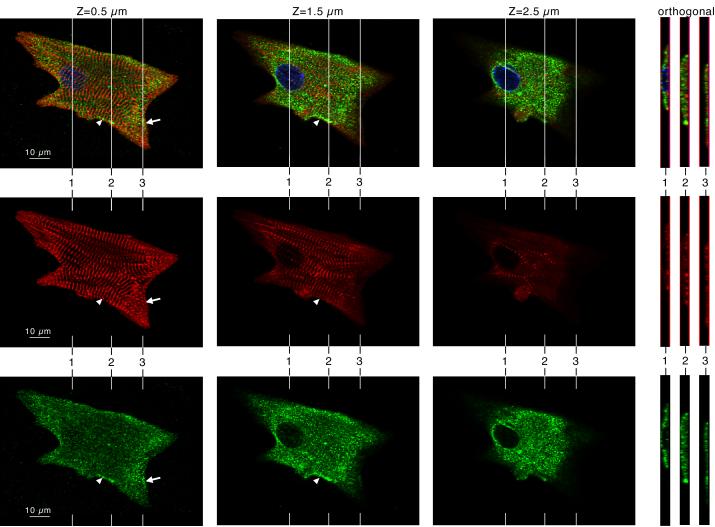
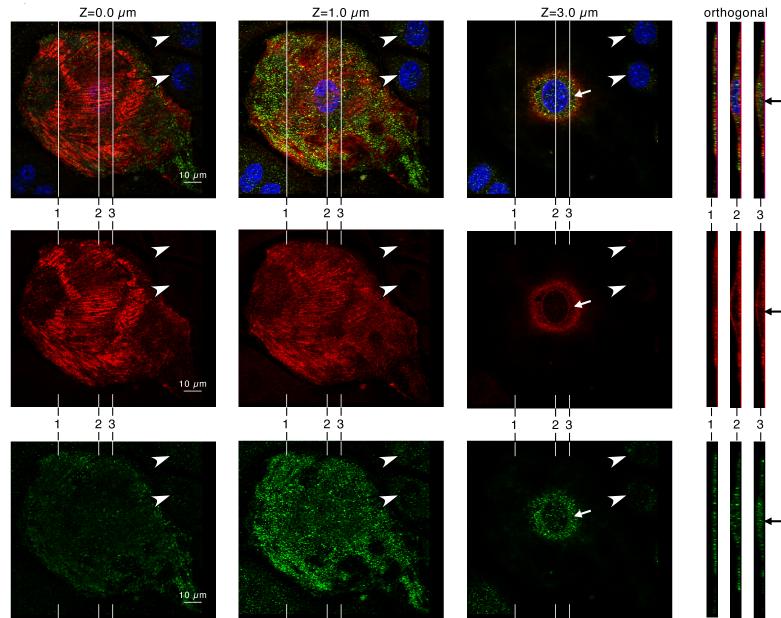
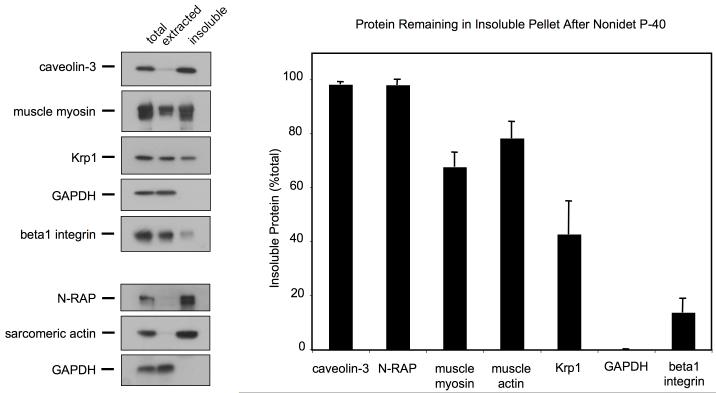
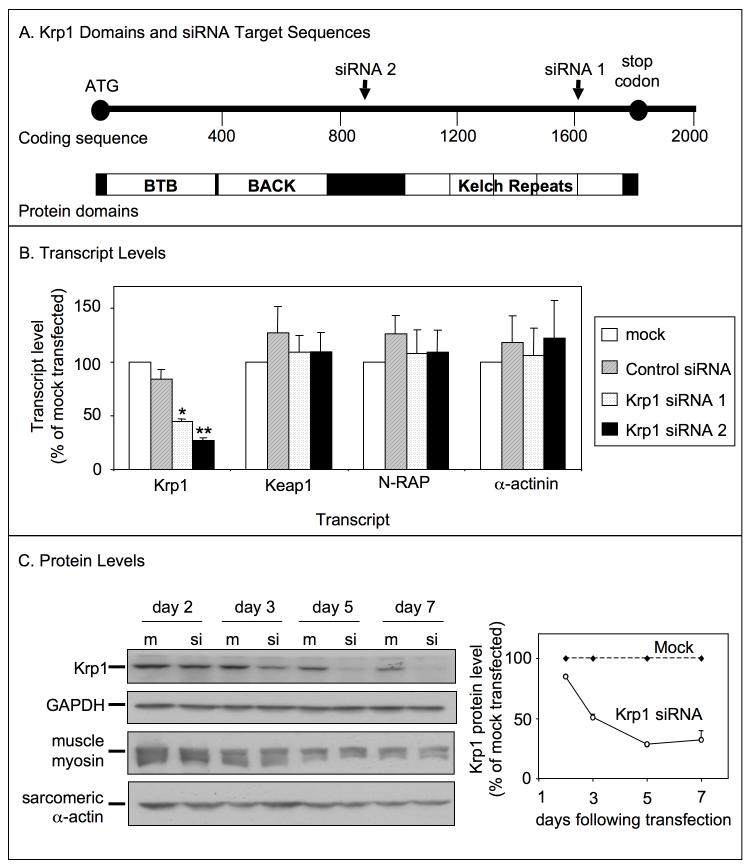
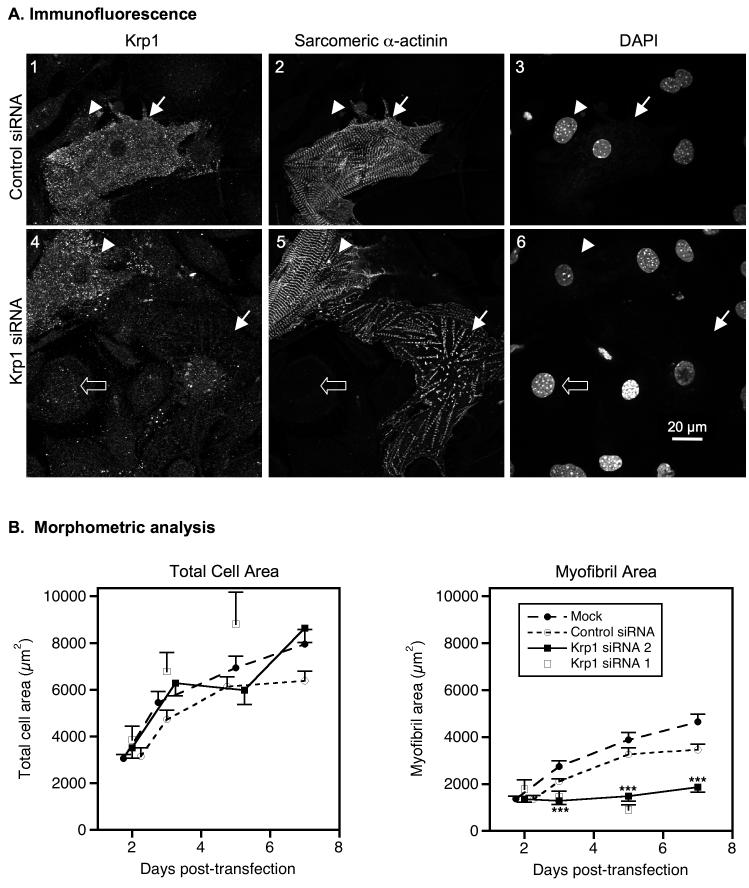
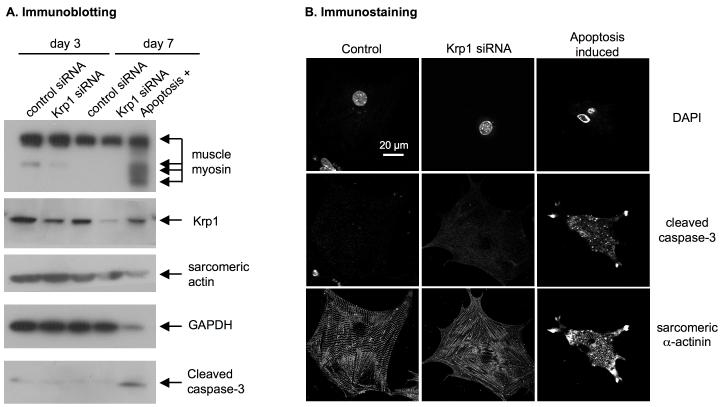
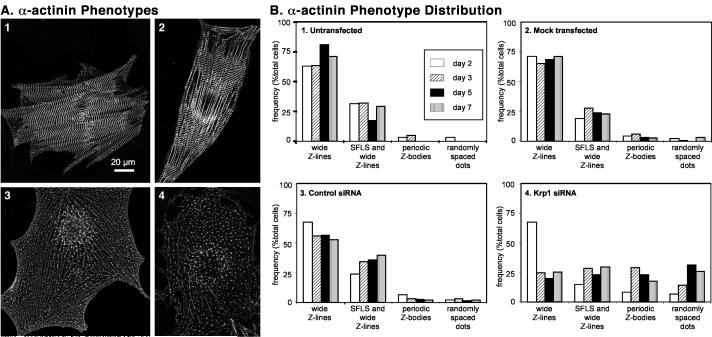
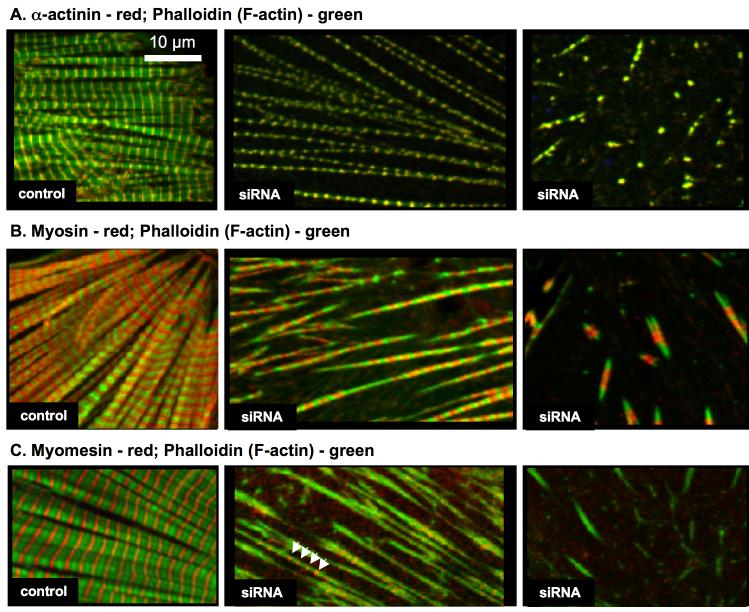
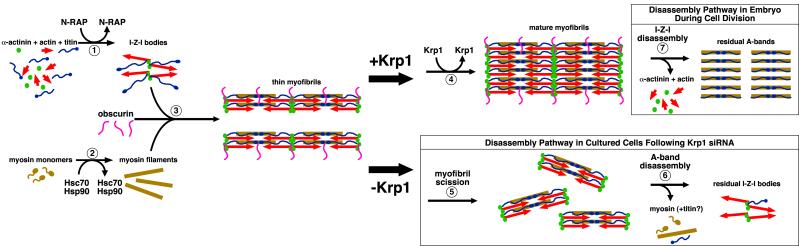
Krp1, also called sarcosin, is a cardiac and skeletal muscle kelch repeat protein hypothesized to promote the assembly of myofibrils, the contractile organelles of striated muscles, through interaction with N-RAP and actin. To elucidate its role, endogenous Krp1 was studied in primary embryonic mouse cardiomyocytes. While immunofluorescence showed punctate Krp1 distribution throughout the cell, detergent extraction revealed a significant pool of Krp1 associated with cytoskeletal elements. Reduction of Krp1 expression with siRNA resulted in specific inhibition of myofibril accumulation with no effect on cell spreading. Immunostaining analysis and electron microscopy revealed that cardiomyocytes lacking Krp1 contained sarcomeric proteins with longitudinal periodicities similar to mature myofibrils, but fibrils remained thin and separated. These thin myofibrils were degraded by a scission mechanism distinct from the myofibril disassembly pathway observed during cell division in the developing heart. The data are consistent with a model in which Krp1 promotes lateral fusion of adjacent thin fibrils into mature, wide myofibrils and contribute insight into mechanisms of myofibrillogenesis and disassembly.
Figures

Similar articles
-
New N-RAP-binding partners alpha-actinin, filamin and Krp1 detected by yeast two-hybrid screening: implications for myofibril assembly.J Cell Sci. 2003 Jun 1;116(Pt 11):2169-78. doi: 10.1242/jcs.00425. Epub 2003 Apr 8. J Cell Sci. 2003. PMID: 12692149
-
Early incorporation of obscurin into nascent sarcomeres: implication for myofibril assembly during cardiac myogenesis.Histochem Cell Biol. 2008 Apr;129(4):463-78. doi: 10.1007/s00418-008-0378-y. Epub 2008 Jan 25. Histochem Cell Biol. 2008. PMID: 18219491 Free PMC article.
-
Role of nonmuscle myosin IIB and N-RAP in cell spreading and myofibril assembly in primary mouse cardiomyocytes.Cell Motil Cytoskeleton. 2008 Sep;65(9):747-61. doi: 10.1002/cm.20299. Cell Motil Cytoskeleton. 2008. PMID: 18615632 Free PMC article.
-
A periodic cytoskeletal lattice in striated muscle.Cell Muscle Motil. 1985;6:287-313. doi: 10.1007/978-1-4757-4723-2_9. Cell Muscle Motil. 1985. PMID: 3888376 Review.
-
Assembly and Maintenance of Myofibrils in Striated Muscle.Handb Exp Pharmacol. 2017;235:39-75. doi: 10.1007/164_2016_53. Handb Exp Pharmacol. 2017. PMID: 27832381 Review.
Cited by
-
Characterization and Comparative Transcriptomic Analysis of Skeletal Muscle in Pekin Duck at Different Growth Stages Using RNA-Seq.Animals (Basel). 2021 Mar 16;11(3):834. doi: 10.3390/ani11030834. Animals (Basel). 2021. PMID: 33809502 Free PMC article.
-
Essential role of the zinc finger transcription factor Casz1 for mammalian cardiac morphogenesis and development.J Biol Chem. 2014 Oct 24;289(43):29801-16. doi: 10.1074/jbc.M114.570416. Epub 2014 Sep 4. J Biol Chem. 2014. PMID: 25190801 Free PMC article.
-
Assembly and Maintenance of Sarcomere Thin Filaments and Associated Diseases.Int J Mol Sci. 2020 Jan 15;21(2):542. doi: 10.3390/ijms21020542. Int J Mol Sci. 2020. PMID: 31952119 Free PMC article. Review.
-
Novel KLHL26 variant associated with a familial case of Ebstein's anomaly and left ventricular noncompaction.Mol Genet Genomic Med. 2020 Apr;8(4):e1152. doi: 10.1002/mgg3.1152. Epub 2020 Jan 27. Mol Genet Genomic Med. 2020. PMID: 31985165 Free PMC article.
-
Kelch proteins: emerging roles in skeletal muscle development and diseases.Skelet Muscle. 2014 Jun 1;4:11. doi: 10.1186/2044-5040-4-11. eCollection 2014. Skelet Muscle. 2014. PMID: 24959344 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
