

The actions of calcium on hair bundle mechanics in mammalian cochlear hair cells
- PMID: 18178649
- PMCID: PMC2267152
- DOI: 10.1529/biophysj.107.123257
The actions of calcium on hair bundle mechanics in mammalian cochlear hair cells
Abstract
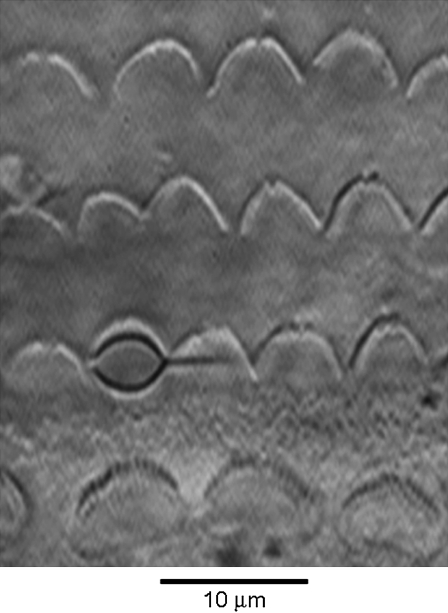
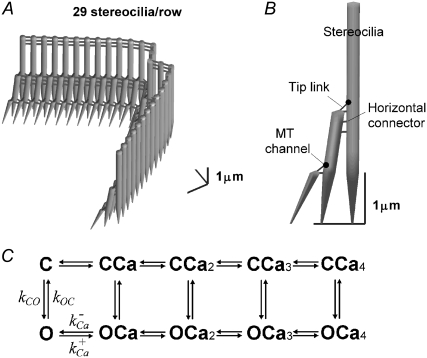
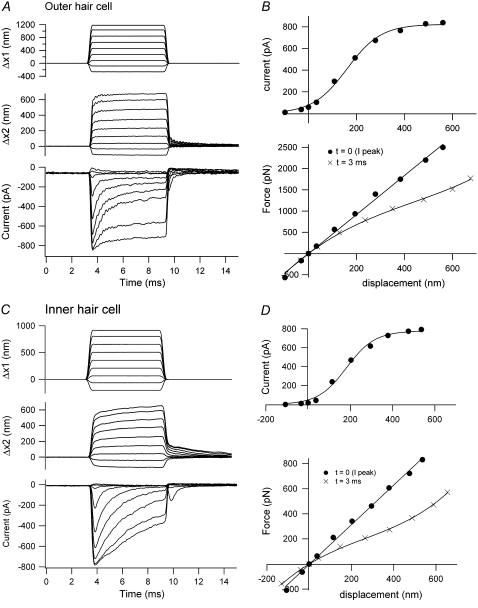
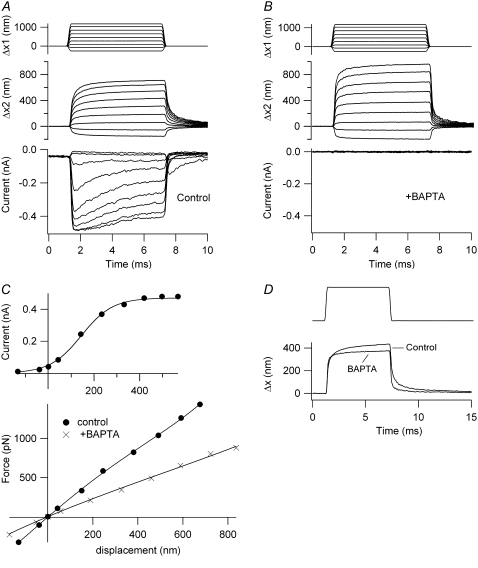
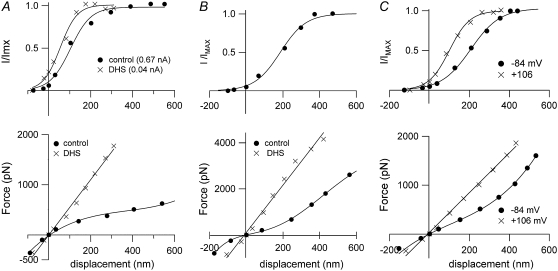
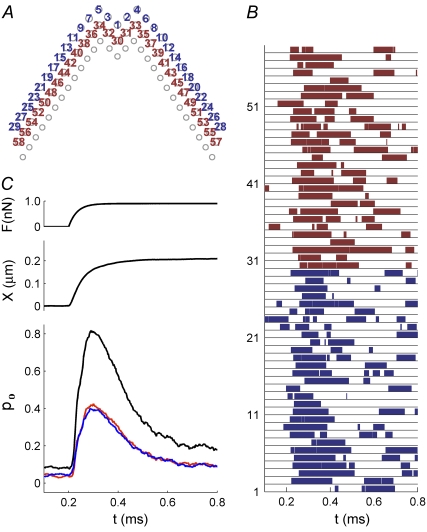
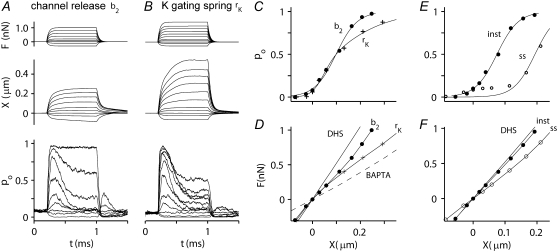
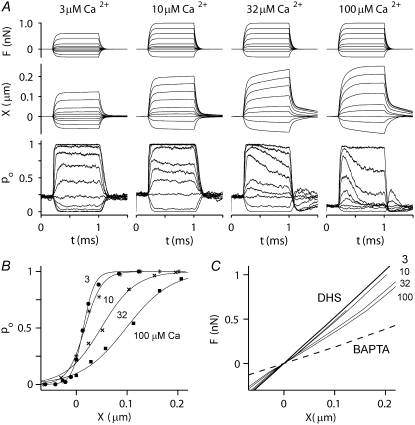
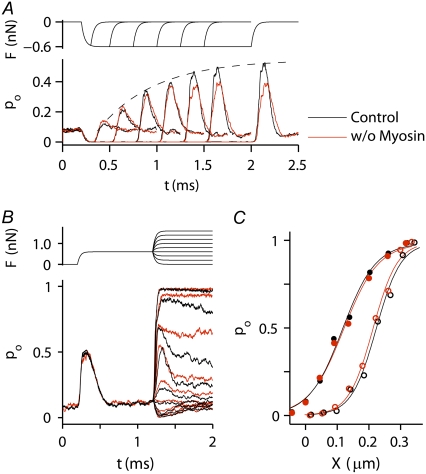
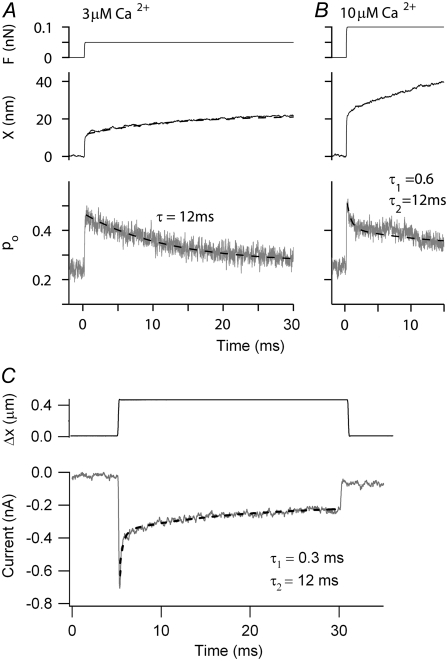
Sound stimuli excite cochlear hair cells by vibration of each hair bundle, which opens mechanotransducer (MT) channels. We have measured hair-bundle mechanics in isolated rat cochleas by stimulation with flexible glass fibers and simultaneous recording of the MT current. Both inner and outer hair-cell bundles exhibited force-displacement relationships with a nonlinearity that reflects a time-dependent reduction in stiffness. The nonlinearity was abolished, and hair-bundle stiffness increased, by maneuvers that diminished calcium influx through the MT channels: lowering extracellular calcium, blocking the MT current with dihydrostreptomycin, or depolarizing to positive potentials. To simulate the effects of Ca(2+), we constructed a finite-element model of the outer hair cell bundle that incorporates the gating-spring hypothesis for MT channel activation. Four calcium ions were assumed to bind to the MT channel, making it harder to open, and, in addition, Ca(2+) was posited to cause either a channel release or a decrease in the gating-spring stiffness. Both mechanisms produced Ca(2+) effects on adaptation and bundle mechanics comparable to those measured experimentally. We suggest that fast adaptation and force generation by the hair bundle may stem from the action of Ca(2+) on the channel complex and do not necessarily require the direct involvement of a myosin motor. The significance of these results for cochlear transduction and amplification are discussed.
Figures

References
-
- Pickles, J. O., S. D. Comis, and M. P. Osborne. 1984. Cross-links between stereocilia in the guinea pig organ of Corti, and their possible relation to sensory transduction. Hear. Res. 15:103–112. - PubMed
-
- Hudspeth, A. J. 1989. How the ear's works work. Nature. 341:397–404. - PubMed
-
- Eatock, R. A. 2000. Adaptation in hair cells. Annu. Rev. Neurosci. 23:285–314. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
