

A class II histone deacetylase acts on newly synthesized histones in Tetrahymena
- PMID: 18178773
- PMCID: PMC2268513
- DOI: 10.1128/EC.00409-07
A class II histone deacetylase acts on newly synthesized histones in Tetrahymena
Abstract
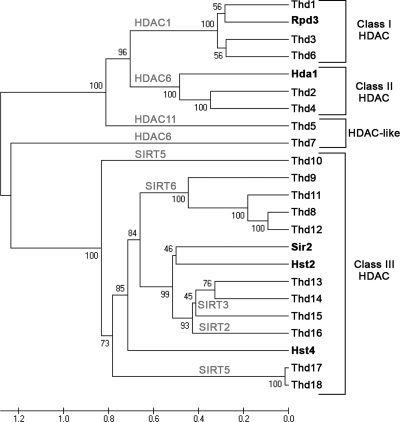
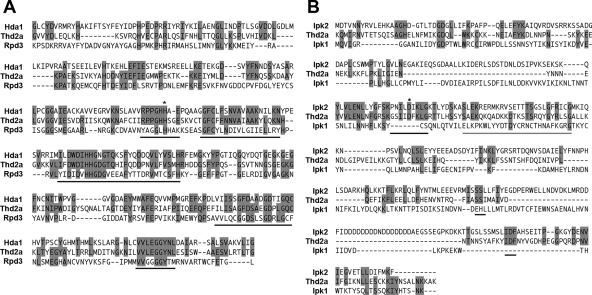
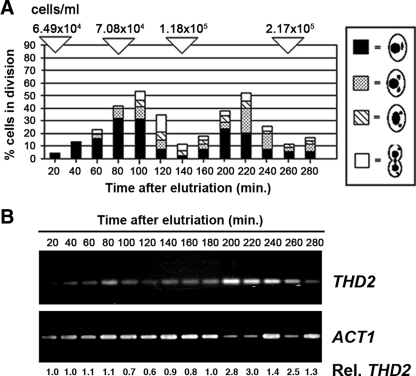
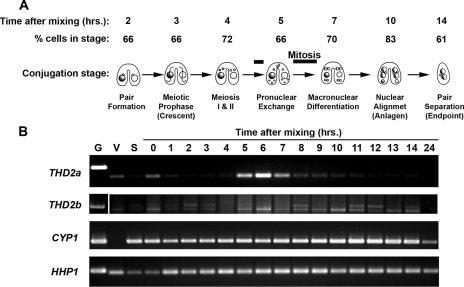
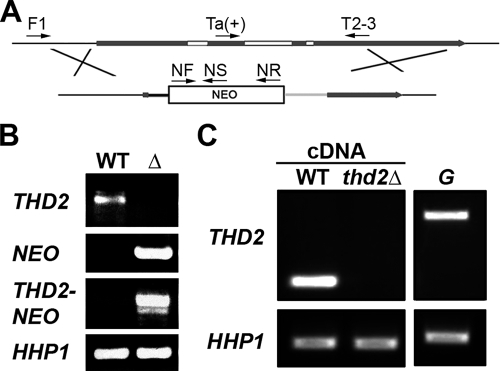
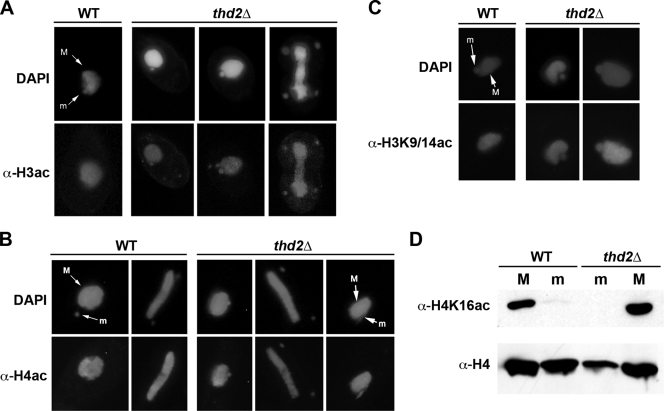
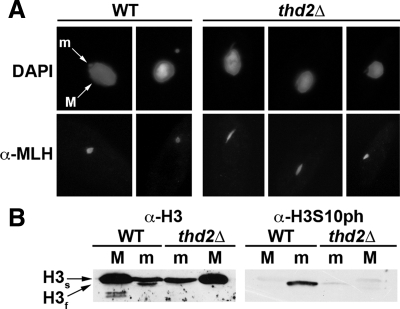
Newly synthesized histones are acetylated prior to their deposition into nucleosomes. Following nucleosome formation and positioning, they are rapidly deacetylated, an event that coincides with further maturation of the chromatin fiber. The histone deacetylases (HDACs) used for histone deposition and de novo chromatin formation are poorly understood. In the ciliate Tetrahymena thermophila, transcription-related deacetylation in the macronucleus is physically separated from deposition-related deacetylation in the micronucleus. This feature was utilized to identify an HDAC named Thd2, a class II HDAC that acts on newly synthesized histones to remove deposition-related acetyl moieties. The THD2 transcript is alternatively spliced, and the major form contains a putative inositol polyphosphate kinase (IPK) domain similar to Ipk2, an enzyme that promotes chromatin remodeling by SWI/SNF remodeling complexes. Cells lacking Thd2, which retain deposition-related acetyl moieties on new histones, exhibit chromatin and cytological phenotypes indicative of a role for Thd2 in chromatin maturation, including the proteolytic processing of histone H3.
Figures


References
-
- Ai, X., and M. R. Parthun. 2004. The nuclear Hat1p/Hat2p complex: a molecular link between type B histone acetyltransferases and chromatin assembly. Mol. Cell 14195-205. - PubMed
-
- Allis, C. D., and M. A. Gorovsky. 1981. Histone phosphorylation in macro- and micronuclei of Tetrahymena thermophila. Biochemistry 203828-3833. - PubMed
-
- Allis, C. D., and J. C. Wiggins. 1984. Histone rearrangements accompany nuclear differentiation and dedifferentiation in Tetrahymena. Dev. Biol. 101282-294. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
