

Cysteines flanking the internal fusion peptide are required for the avian sarcoma/leukosis virus glycoprotein to mediate the lipid mixing stage of fusion with high efficiency
- PMID: 18184714
- PMCID: PMC2259008
- DOI: 10.1128/JVI.02266-07
Cysteines flanking the internal fusion peptide are required for the avian sarcoma/leukosis virus glycoprotein to mediate the lipid mixing stage of fusion with high efficiency
Abstract
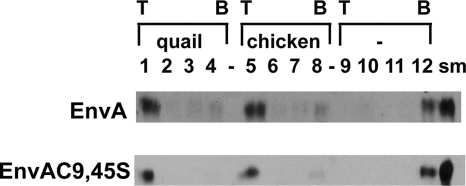
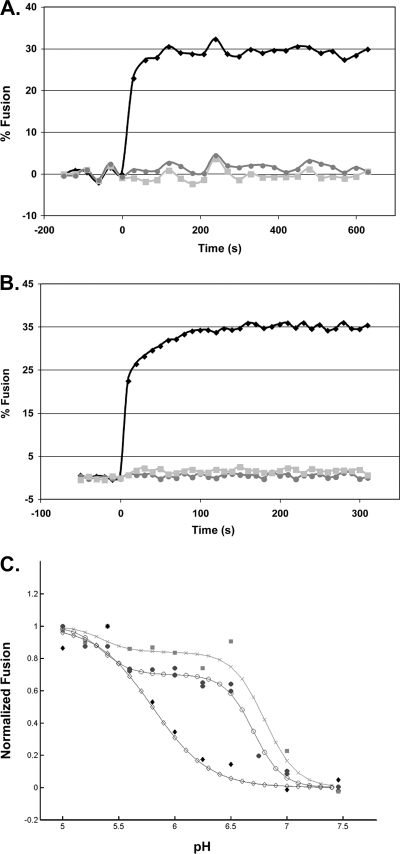
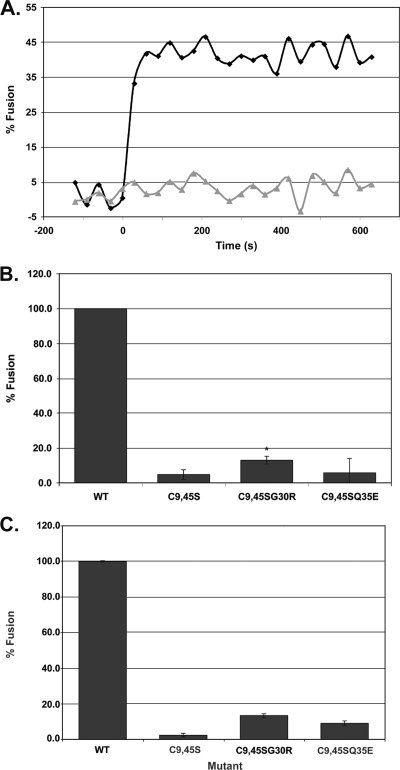
We previously showed that the cysteines flanking the internal fusion peptide of the avian sarcoma/leukosis virus subtype A (ASLV-A) Env (EnvA) are important for infectivity and cell-cell fusion. Here we define the stage of fusion at which the cysteines are required. The flanking cysteines are dispensable for receptor-triggered membrane association but are required for the lipid mixing step of fusion, which, interestingly, displays a high pH onset and a biphasic profile. Second-site mutations that partially restore infection partially restore lipid mixing. These findings indicate that the cysteines flanking the internal fusion peptide of EnvA (and perhaps by analogy Ebola virus glycoprotein) are important for the foldback stage of the conformational changes that lead to membrane merger.
Figures
References
-
- Baker, K. A., R. E. Dutch, R. A. Lamb, and T. S. Jardetzky. 1999. Structural basis for paramyxovirus-mediated membrane fusion. Mol. Cell 3309-319. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
