

Unregulated cytosolic dopamine causes neurodegeneration associated with oxidative stress in mice
- PMID: 18184785
- PMCID: PMC6670521
- DOI: 10.1523/JNEUROSCI.3602-07.2008
Unregulated cytosolic dopamine causes neurodegeneration associated with oxidative stress in mice
Abstract
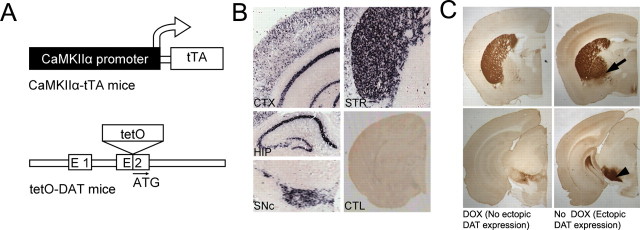
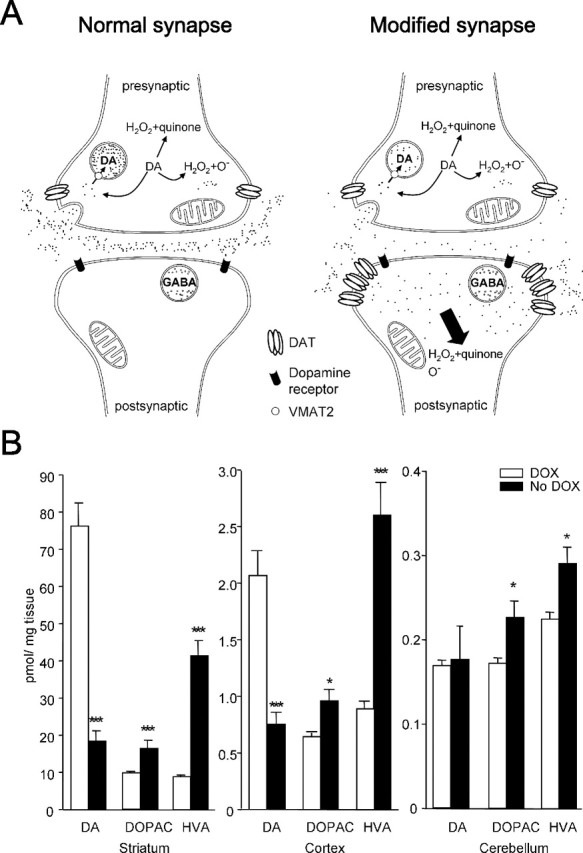
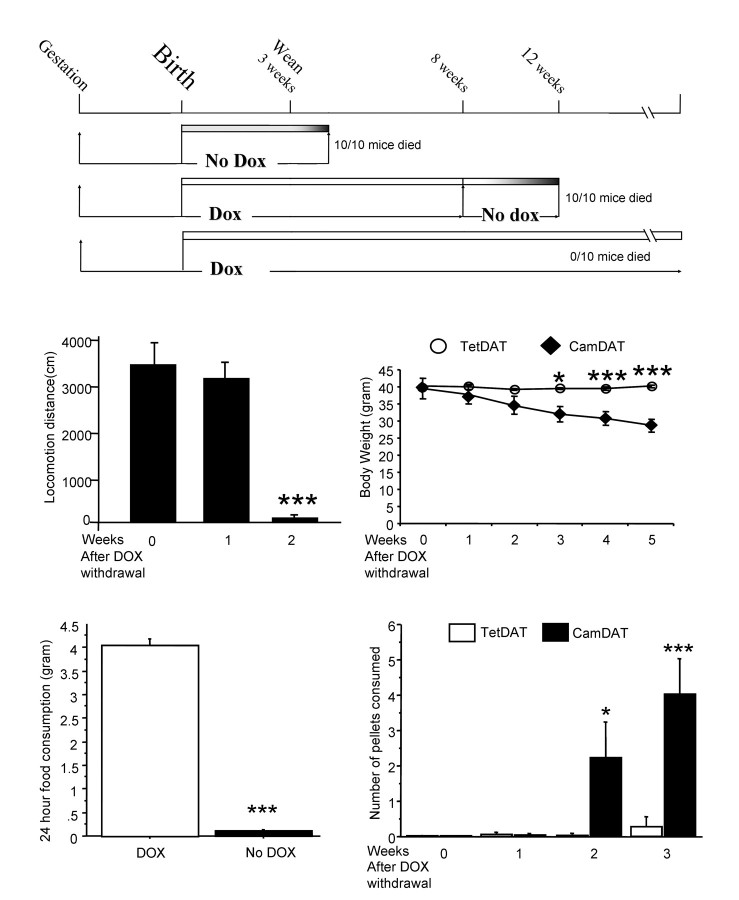
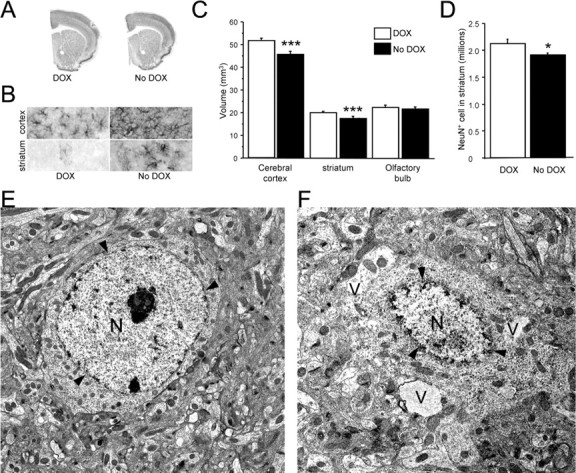
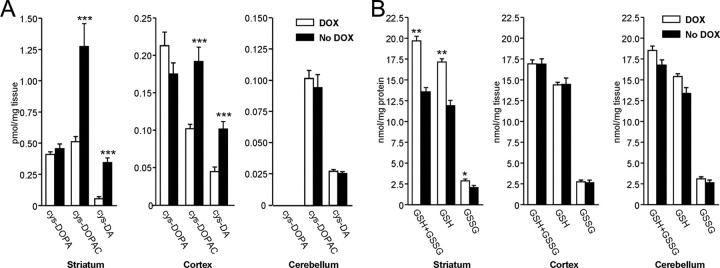
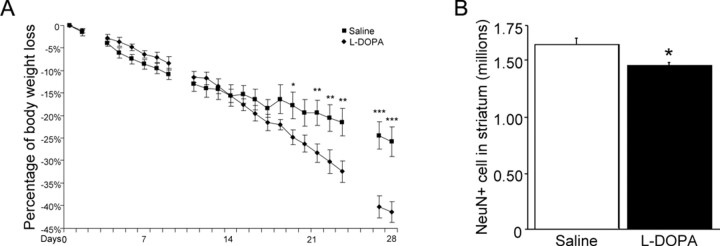
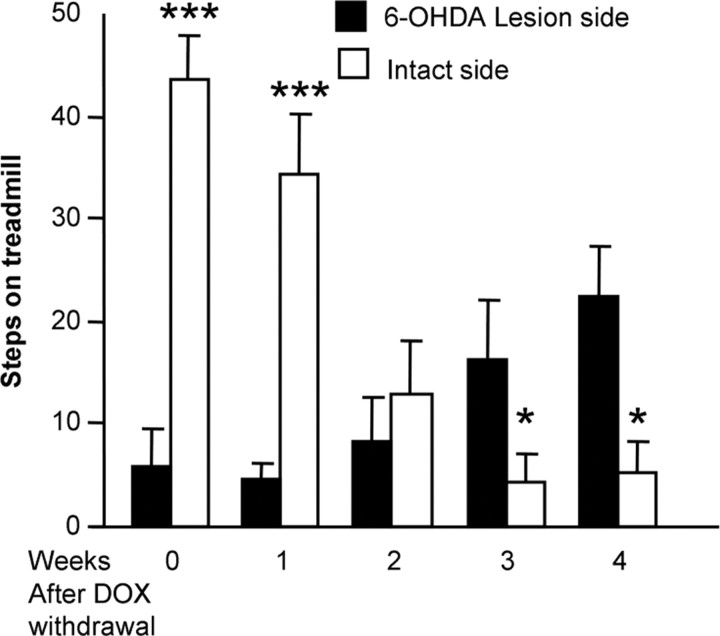
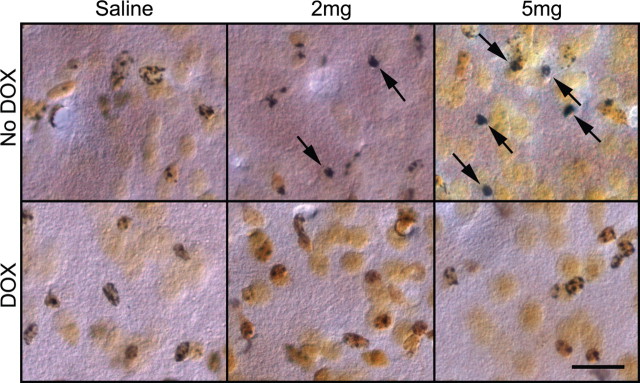
The role of dopamine as a vulnerability factor and a toxic agent in Parkinson's disease (PD) is still controversial, yet the presumed dopamine toxicity is partly responsible for the "DOPA-sparing" clinical practice that avoids using L-3,4-dihydroxyphenylalanine (L-DOPA), a dopamine precursor, in early PD. There is a lack of studies on animal models that directly isolate dopamine as one determining factor in causing neurodegeneration. To address this, we have generated a novel transgenic mouse model in which striatal neurons are engineered to take up extracellular dopamine without acquiring regulatory mechanisms found in dopamine neurons. These mice developed motor dysfunctions and progressive neurodegeneration in the striatum within weeks. The neurodegeneration was accompanied by oxidative stress, evidenced by substantial oxidative protein modifications and decrease in glutathione. Ultrastructural morphologies of degenerative cells suggest necrotic neurodegeneration. Moreover, L-DOPA accelerated neurodegeneration and worsened motor dysfunction. In contrast, reducing dopamine input to striatum by lesioning the medial forebrain bundle attenuated motor dysfunction. These data suggest that pathology in genetically modified striatal neurons depends on their dopamine supply. These neurons were also supersensitive to neurotoxin. A very low dose of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (5 mg/kg) caused profound neurodegeneration of striatal neurons, but not midbrain dopamine neurons. Our results provide the first in vivo evidence that chronic exposure to unregulated cytosolic dopamine alone is sufficient to cause neurodegeneration. The present study has significant clinical implications, because dopamine replacement therapy is the mainstay of PD treatment. In addition, our model provides an efficient in vivo approach to test therapeutic agents for PD.
Figures
References
-
- Agid Y, Ahlskog E, Albanese A, Calne D, Chase T, De Yebenes J, Factor S, Fahn S, Gershanik O, Goetz C, Koller W, Kurth M, Lang A, Lees A, Lewitt P, Marsden D, Melamed E, Michel PP, Mizuno Y, Obeso J, et al. Levodopa in the treatment of Parkinson's disease: a consensus meeting. Mov Disord. 1999;14:911–913. - PubMed
-
- Barzilai A, Daily D, Zilkha-Falb R, Ziv I, Offen D, Melamed E, Shirvan A. The molecular mechanisms of dopamine toxicity. Adv Neurol. 2003;91:73–82. - PubMed
-
- Bonifati V, Rizzu P, van Baren MJ, Schaap O, Breedveld GJ, Krieger E, Dekker MC, Squitieri F, Ibanez P, Joosse M, van Dongen JW, Vanacore N, van Swieten JC, Brice A, Meco G, van Duijn CM, Oostra BA, Heutink P. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science. 2003;299:256–259. - PubMed
-
- Cappai R, Leck SL, Tew DJ, Williamson NA, Smith DP, Galatis D, Sharples RA, Curtain CC, Ali FE, Cherny RA, Culvenor JG, Bottomley SP, Masters CL, Barnham KJ, Hill AF. Dopamine promotes alpha-synuclein aggregation into SDS-resistant soluble oligomers via a distinct folding pathway. FASEB J. 2005;19:1377–1379. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases