

The consequences of response nonlinearities for interpretation of spectrotemporal receptive fields
- PMID: 18184787
- PMCID: PMC6670552
- DOI: 10.1523/JNEUROSCI.1775-07.2007
The consequences of response nonlinearities for interpretation of spectrotemporal receptive fields
Abstract
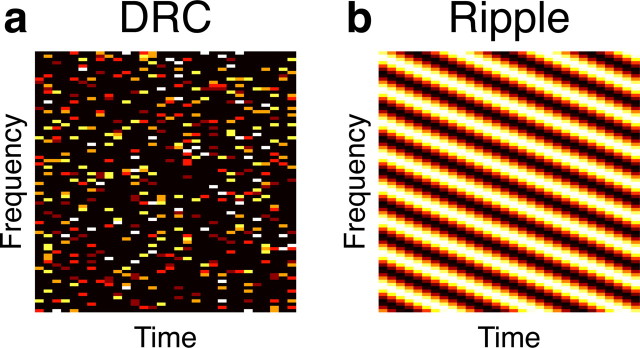
Neurons in the central auditory system are often described by the spectrotemporal receptive field (STRF), conventionally defined as the best linear fit between the spectrogram of a sound and the spike rate it evokes. An STRF is often assumed to provide an estimate of the receptive field of a neuron, i.e., the spectral and temporal range of stimuli that affect the response. However, when the true stimulus-response function is nonlinear, the STRF will be stimulus dependent, and changes in the stimulus properties can alter estimates of the sign and spectrotemporal extent of receptive field components. We demonstrate analytically and in simulations that, even when uncorrelated stimuli are used, interactions between simple neuronal nonlinearities and higher-order structure in the stimulus can produce STRFs that show contributions from time-frequency combinations to which the neuron is actually insensitive. Only when spectrotemporally independent stimuli are used does the STRF reliably indicate features of the underlying receptive field, and even then it provides only a conservative estimate. One consequence of these observations, illustrated using natural stimuli, is that a stimulus-induced change in an STRF could arise from a consistent but nonlinear neuronal response to stimulus ensembles with differing higher-order dependencies. Thus, although the responses of higher auditory neurons may well involve adaptation to the statistics of different stimulus ensembles, stimulus dependence of STRFs alone, or indeed of any overly constrained stimulus-response mapping, cannot demonstrate the nature or magnitude of such effects.
Figures









References
-
- Aertsen AM, Johannesma PI. A comparison of the spectro-temporal sensitivity of auditory neurons to tonal and natural stimuli. Biol Cybern. 1981;42:145–156. - PubMed
-
- Aertsen AM, Olders JH, Johannesma PI. Spectro-temporal receptive fields of auditory neurons in the grassfrog. III. Analysis of the stimulus-event relation for natural stimuli. Biol Cybern. 1981;39:195–209. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources