

Glutamate receptor-dependent increments in lactate, glucose and oxygen metabolism evoked in rat cerebellum in vivo
- PMID: 18187464
- PMCID: PMC2375663
- DOI: 10.1113/jphysiol.2007.144154
Glutamate receptor-dependent increments in lactate, glucose and oxygen metabolism evoked in rat cerebellum in vivo
Abstract
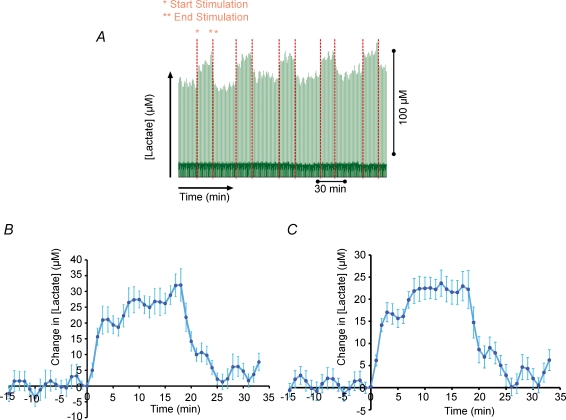
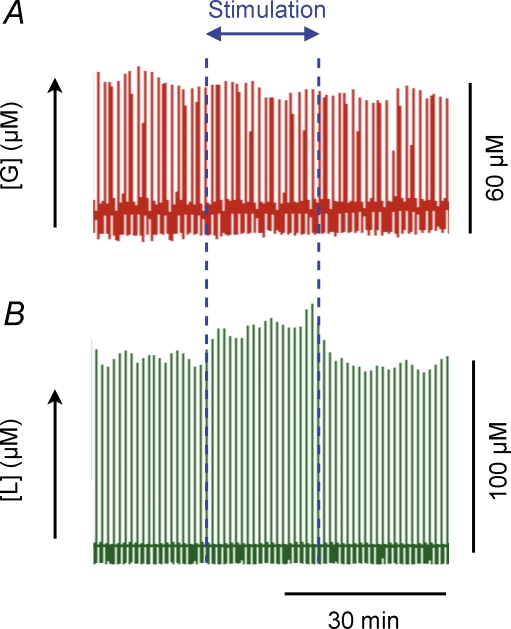
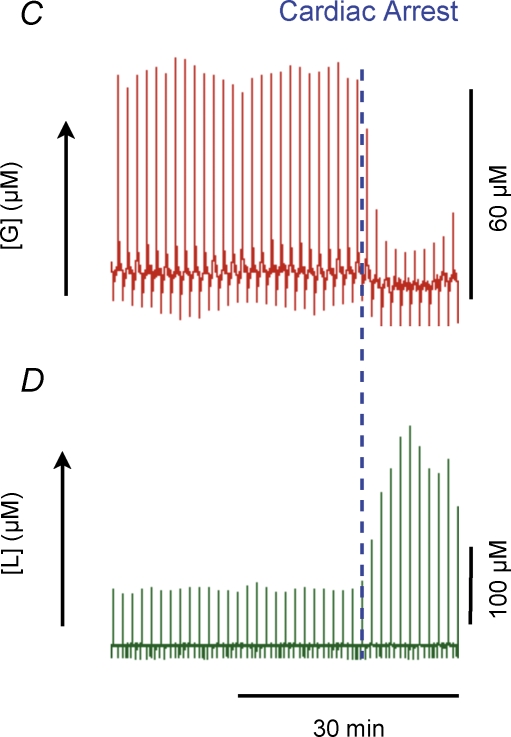
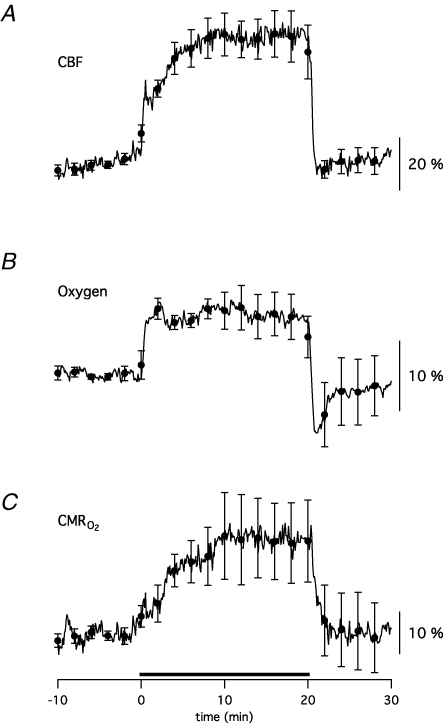
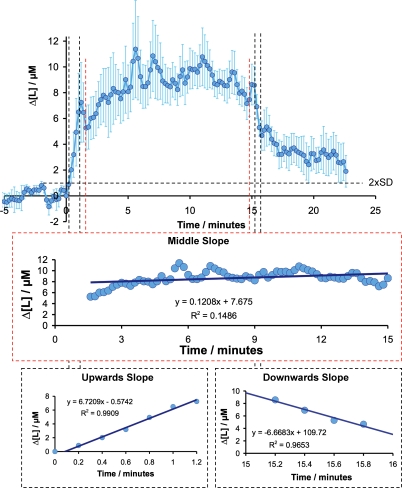
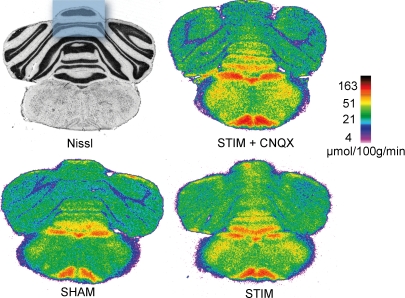
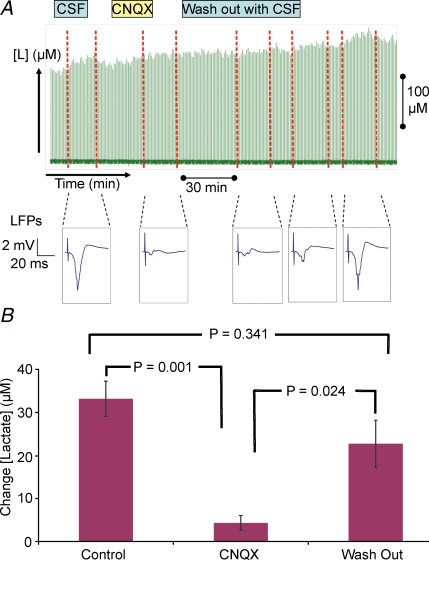
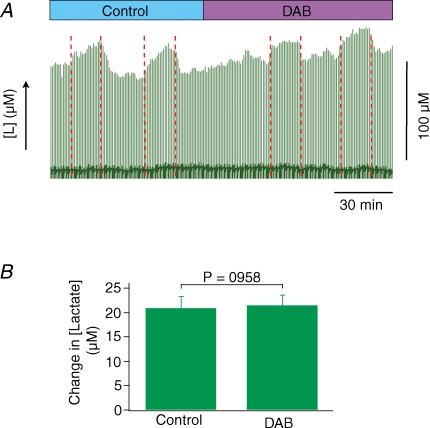
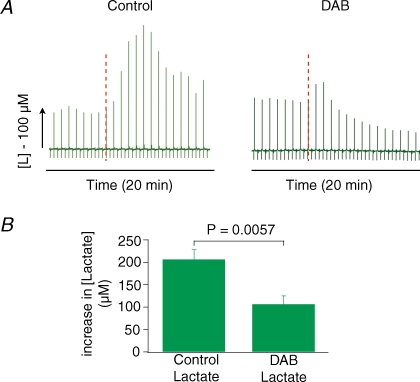
Neuronal activity is tightly coupled with brain energy metabolism. Numerous studies have suggested that lactate is equally important as an energy substrate for neurons as glucose. Lactate production is reportedly triggered by glutamate uptake, and independent of glutamate receptor activation. Here we show that climbing fibre stimulation of cerebellar Purkinje cells increased extracellular lactate by 30% within 30 s of stimulation, but not for briefer stimulation periods. To explore whether lactate production was controlled by pre- or postsynaptic events we silenced AMPA receptors with CNQX. This blocked all evoked rises in postsynaptic activity, blood flow, and glucose and oxygen consumption. CNQX also abolished rises in lactate concomitantly with marked reduction in postsynaptic currents. Rises in lactate were unaffected by inhibition of glycogen phosphorylase, suggesting that lactate production was independent of glycogen breakdown. Stimulated lactate production in cerebellum is derived directly from glucose uptake, and coupled to neuronal activity via AMPA receptor activation.
Figures
Comment in
-
A new pathway for lactate production in the CNS.J Physiol. 2008 Mar 1;586(5):1207-8. doi: 10.1113/jphysiol.2008.151373. J Physiol. 2008. PMID: 18310130 Free PMC article. No abstract available.
Similar articles
-
A new pathway for lactate production in the CNS.J Physiol. 2008 Mar 1;586(5):1207-8. doi: 10.1113/jphysiol.2008.151373. J Physiol. 2008. PMID: 18310130 Free PMC article. No abstract available.
-
Differential effects of NMDA and AMPA/kainate receptor antagonists on nitric oxide production in rat brain following intrahippocampal injection.Brain Res Bull. 2005 Sep 30;67(1-2):133-41. doi: 10.1016/j.brainresbull.2005.06.019. Brain Res Bull. 2005. PMID: 16140172
-
Modification of activity-dependent increases in cerebellar blood flow by extracellular potassium in anaesthetized rats.J Physiol. 1999 Oct 1;520 Pt 1(Pt 1):281-92. doi: 10.1111/j.1469-7793.1999.00281.x. J Physiol. 1999. PMID: 10517819 Free PMC article.
-
Synaptic activation of AMPA receptors inhibits GABA release from cerebellar interneurons.Nat Neurosci. 2000 Jun;3(6):551-8. doi: 10.1038/75718. Nat Neurosci. 2000. PMID: 10816310
-
Ampakine CX546 bolsters energetic response of astrocytes: a novel target for cognitive-enhancing drugs acting as alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptor modulators.J Neurochem. 2005 Feb;92(3):668-77. doi: 10.1111/j.1471-4159.2004.02905.x. J Neurochem. 2005. PMID: 15659236
Cited by
-
Multifunctional role of astrocytes as gatekeepers of neuronal energy supply.Front Cell Neurosci. 2013 Apr 10;7:38. doi: 10.3389/fncel.2013.00038. eCollection 2013. Front Cell Neurosci. 2013. PMID: 23596393 Free PMC article.
-
Calcium signaling in brain mitochondria: interplay of malate aspartate NADH shuttle and calcium uniporter/mitochondrial dehydrogenase pathways.J Biol Chem. 2009 Mar 13;284(11):7091-9. doi: 10.1074/jbc.M808066200. Epub 2009 Jan 7. J Biol Chem. 2009. PMID: 19129175 Free PMC article.
-
Global and Regional Derangements of Cerebral Blood Flow and Diffusion Magnetic Resonance Imaging after Pediatric Cardiac Arrest.J Pediatr. 2016 Feb;169:28-35.e1. doi: 10.1016/j.jpeds.2015.10.003. Epub 2015 Nov 11. J Pediatr. 2016. PMID: 26561380 Free PMC article.
-
Interplay between Energy Supply and Glutamate Toxicity in the Primary Cortical Culture.Biomolecules. 2024 Apr 30;14(5):543. doi: 10.3390/biom14050543. Biomolecules. 2024. PMID: 38785950 Free PMC article.
-
Spreading Depression, Spreading Depolarizations, and the Cerebral Vasculature.Physiol Rev. 2015 Jul;95(3):953-93. doi: 10.1152/physrev.00027.2014. Physiol Rev. 2015. PMID: 26133935 Free PMC article. Review.
References
-
- Bergersen L, Waerhaug O, Helm J, Thomas M, Laake P, Davies AJ, Wilson MC, Halestrap AP, Ottersen OP. A novel postsynaptic density protein: the monocarboxylate transporter MCT2 is co-localized with δ-glutamate receptors in postsynaptic densities of parallel fiber-Purkinje cell synapses. Exp Brain Res. 2001;136:523–534. - PubMed
-
- Bergersen LH, Magistretti PJ, Pellerin L. Selective postsynaptic co-localization of MCT2 with AMPA receptor GluR2/3 subunits at excitatory synapses exhibiting AMPA receptor trafficking. Cerebr Cortex. 2005;15:361–370. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources