

Parathyroid hormone increases activating transcription factor 4 expression and activity in osteoblasts: requirement for osteocalcin gene expression
- PMID: 18187540
- PMCID: PMC2276723
- DOI: 10.1210/en.2007-1573
Parathyroid hormone increases activating transcription factor 4 expression and activity in osteoblasts: requirement for osteocalcin gene expression
Abstract
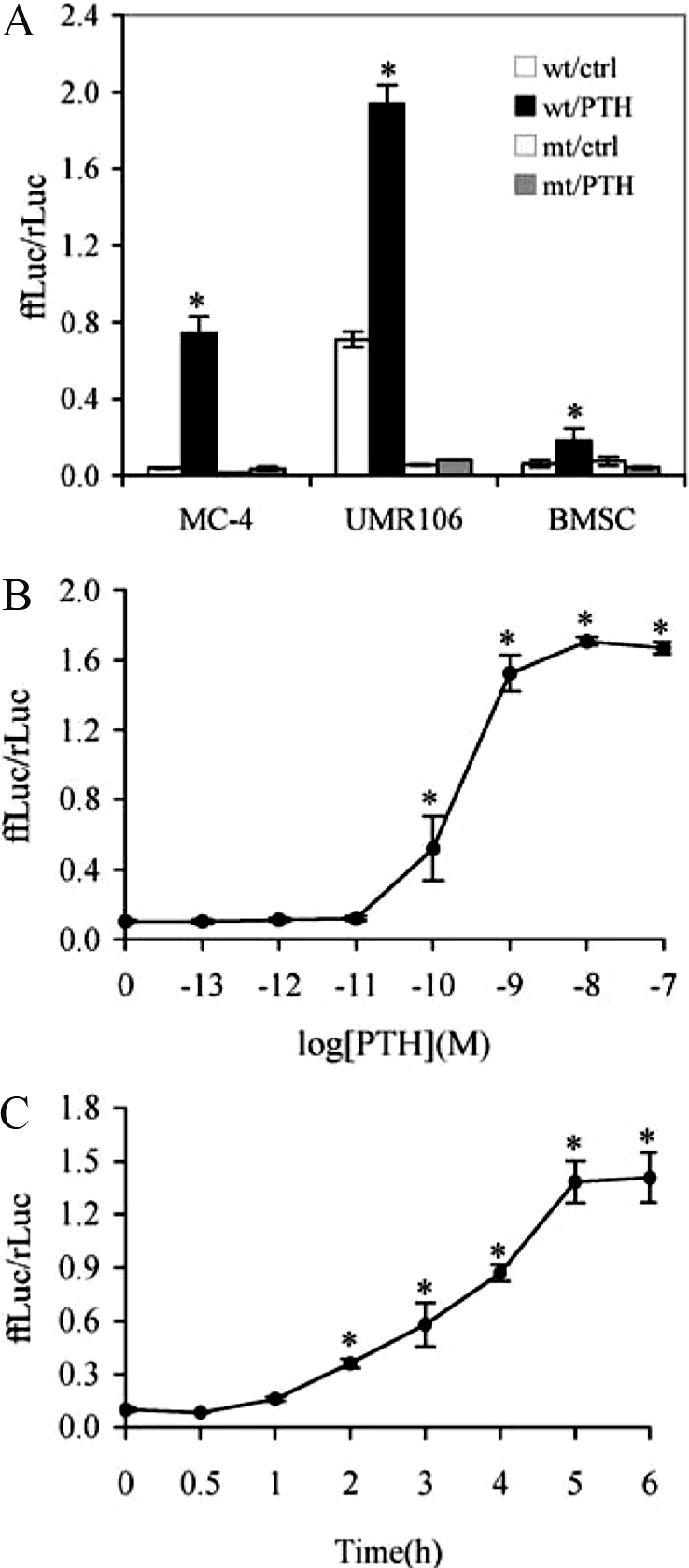
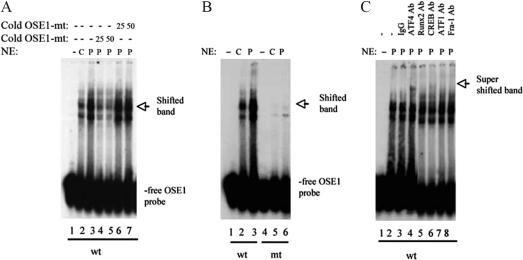
PTH is an important peptide hormone regulator of calcium homeostasis and osteoblast function. However, its mechanism of action in osteoblasts is poorly understood. Our previous study demonstrated that PTH activates mouse osteocalcin (Ocn) gene 2 promoter through the osteoblast-specific element 1 site, a recently identified activating transcription factor-4 (ATF4) -binding element. In the present study, we examined effects of PTH on ATF4 expression and activity as well as the requirement for ATF4 in the regulation of Ocn by PTH. Results show that PTH elevated levels of ATF4 mRNA and protein in a dose- and time-dependent manner. This PTH regulation requires transcriptional activity but not de novo protein synthesis. PTH also increased binding of nuclear extracts to osteoblast-specific element 1 DNA. PTH stimulated ATF4-dependent transcriptional activity mainly through protein kinase A with a lesser requirement for protein kinase C and MAPK/ERK pathways. Lastly, PTH stimulation of Ocn expression was lost by small interfering RNA down-regulation of ATF4 in MC-4 cells and Atf4(-/-) bone marrow stromal cells. Collectively, these studies for the first time demonstrate that PTH increases ATF4 expression and activity and that ATF4 is required for PTH induction of Ocn expression in osteoblasts.
Figures





Similar articles
-
The basic helix loop helix transcription factor Twist1 is a novel regulator of ATF4 in osteoblasts.J Cell Biochem. 2012 Jan;113(1):70-9. doi: 10.1002/jcb.23329. J Cell Biochem. 2012. PMID: 21866569 Free PMC article.
-
Parathyroid hormone induction of the osteocalcin gene. Requirement for an osteoblast-specific element 1 sequence in the promoter and involvement of multiple-signaling pathways.J Biol Chem. 2004 Feb 13;279(7):5329-37. doi: 10.1074/jbc.M311547200. Epub 2003 Nov 21. J Biol Chem. 2004. PMID: 14634012
-
PGC-1alpha is induced by parathyroid hormone and coactivates Nurr1-mediated promoter activity in osteoblasts.Bone. 2006 Nov;39(5):1018-1025. doi: 10.1016/j.bone.2006.04.023. Epub 2006 Jun 12. Bone. 2006. PMID: 16765661
-
Parathyroid hormone-dependent signaling pathways regulating genes in bone cells.Gene. 2002 Jan 9;282(1-2):1-17. doi: 10.1016/s0378-1119(01)00798-3. Gene. 2002. PMID: 11814673 Review.
-
Advances in the roles of ATF4 in osteoporosis.Biomed Pharmacother. 2023 Dec 31;169:115864. doi: 10.1016/j.biopha.2023.115864. Epub 2023 Nov 9. Biomed Pharmacother. 2023. PMID: 37948991 Review.
Cited by
-
atf4 promotes β-catenin expression and osteoblastic differentiation of bone marrow mesenchymal stem cells.Int J Biol Sci. 2013;9(3):256-66. doi: 10.7150/ijbs.5898. Epub 2013 Feb 27. Int J Biol Sci. 2013. PMID: 23494915 Free PMC article.
-
The basic helix loop helix transcription factor Twist1 is a novel regulator of ATF4 in osteoblasts.J Cell Biochem. 2012 Jan;113(1):70-9. doi: 10.1002/jcb.23329. J Cell Biochem. 2012. PMID: 21866569 Free PMC article.
-
Critical role of AKT protein in myeloma-induced osteoclast formation and osteolysis.J Biol Chem. 2013 Oct 18;288(42):30399-30410. doi: 10.1074/jbc.M113.469973. Epub 2013 Sep 4. J Biol Chem. 2013. PMID: 24005670 Free PMC article. Clinical Trial.
-
Activating transcription factor 4 regulates osteoclast differentiation in mice.J Clin Invest. 2010 Aug;120(8):2755-66. doi: 10.1172/JCI42106. Epub 2010 Jul 12. J Clin Invest. 2010. PMID: 20628199 Free PMC article.
-
Regulation of gene expression in osteoblasts.Biofactors. 2010 Jan-Feb;36(1):25-32. doi: 10.1002/biof.72. Biofactors. 2010. PMID: 20087883 Free PMC article. Review.
References
-
- Neer RM, Arnaud CD, Zanchetta JR, Prince R, Gaich GA, Reginster JY, Hodsman AB, Eriksen EF, Ish-Shalom S, Genant HK, Wang O, Mitlak BH 2001 Effect of parathyroid hormone (1–34) on fractures and bone mineral density in postmenopausal women with osteoporosis. N Engl J Med 344:1434–1441 - PubMed
-
- Demiralp B, Chen HL, Koh AJ, Keller ET, McCauley LK 2002 Anabolic actions of parathyroid hormone during bone growth are dependent on c-fos. Endocrinology 143:4038–4047 - PubMed
-
- Partridge NC, Alcorn D, Michelangeli VP, Kemp BE, Ryan GB, Martin TJ 1981 Functional properties of hormonally responsive cultured normal and malignant rat osteoblastic cells. Endocrinology 108:213–219 - PubMed
-
- McCauley LK, Koh AJ, Beecher CA, Cui Y, Decker JD, Franceschi RT 1995 Effects of differentiation and transforming growth factor β1 on PTH/PTHrP receptor mRNA levels in MC3T3-E1 cells. J Bone Miner Res 10:1243–1255 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
