

Blood cell interactions and segregation in flow
- PMID: 18188702
- PMCID: PMC2752714
- DOI: 10.1007/s10439-007-9429-0
Blood cell interactions and segregation in flow
Abstract
For more than a century, pioneering researchers have been using novel experimental and computational approaches to probe the mysteries of blood flow. Thanks to their efforts, we know that blood cells generally prefer to migrate to the axis of flow, that red and white cells segregate in flow, and that cell deformability and their tendency to reversibly aggregate contribute to the non-Newtonian nature of this unique fluid. All of these properties have beneficial physiological consequences, allowing blood to perform a variety of critical functions. Our current understanding of these unusual flow properties of blood have been made possible by the ingenuity and diligence of a number of researchers, including Harry Goldsmith, who developed novel technologies to visualize and quantify the flow of blood at the level of individual cells. Here we summarize efforts in our lab to continue this tradition and to further our understanding of how blood cells interact with each other and with the blood vessel wall.
Figures

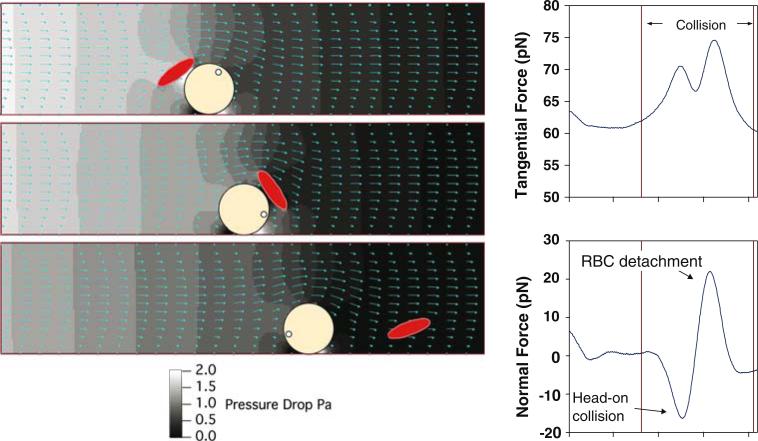
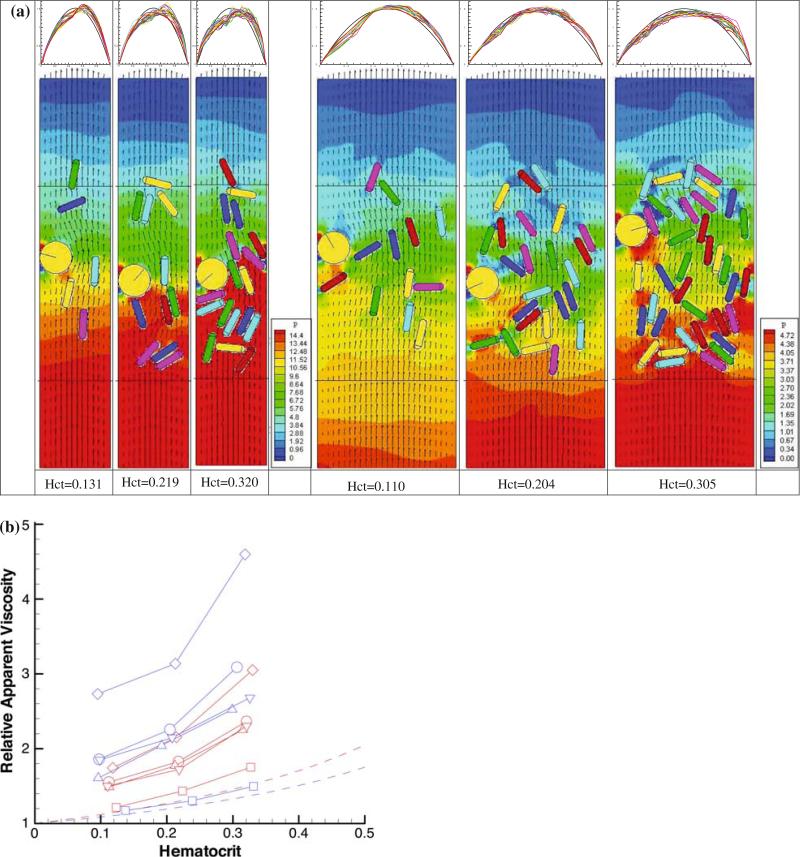

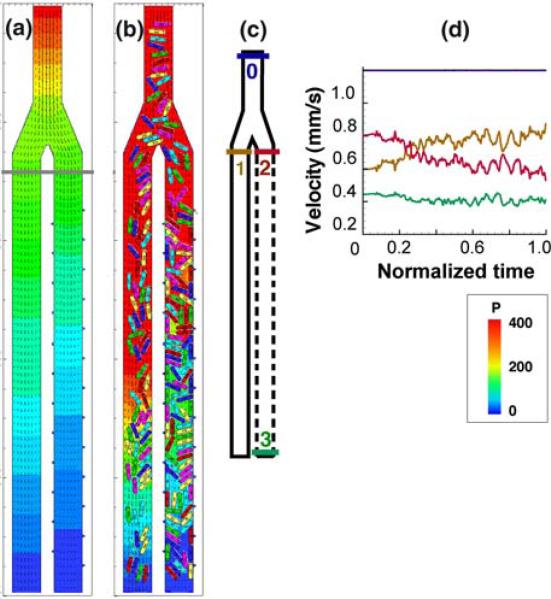

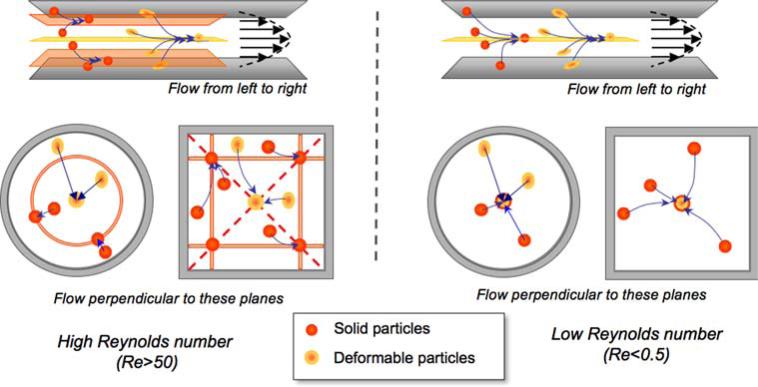

References
-
- Bell GI. Models for the specific adhesion of cells to cells. Science. 1978;200:618–627. - PubMed
-
- Bishop JJ, Nance P, Popel AS, Intaglietta M, Johnson PC. Effect of erythrocyte aggregation on velocity profiles in venules. Am. J. Physiol. Heart Circ. Physiol. 2001;280:H222–H236. - PubMed
-
- Bishop JJ, Popel AS, Intaglietta M, Johnson PC. Effects of erythrocyte aggregation and venous network geometry on red blood cell axial migration. Am. J. Physiol. Heart Circ. Physiol. 2001;281:H939–H950. - PubMed
-
- Brown EB, Campbell RB, Tsuzuki Y, Xu L, Carmeliet P, Fukumura D, Jain RK. In vivo measurement of gene expression, angiogenesis and physiological function in tumors using multiphoton laser scanning microscopy. Nat. Med. 2001;7:864–868. - PubMed
-
- Butcher EC, Lewinsohn D, Duijvestijn A, Bargatze R, Wu N, Jalkanen S. Interactions between endothelial cells and leukocytes. J. Cell. Biochem. 1986;30:121–131. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
