

The spinal nucleus of the bulbocavernosus: firsts in androgen-dependent neural sex differences
- PMID: 18191128
- PMCID: PMC2423220
- DOI: 10.1016/j.yhbeh.2007.11.008
The spinal nucleus of the bulbocavernosus: firsts in androgen-dependent neural sex differences
Abstract
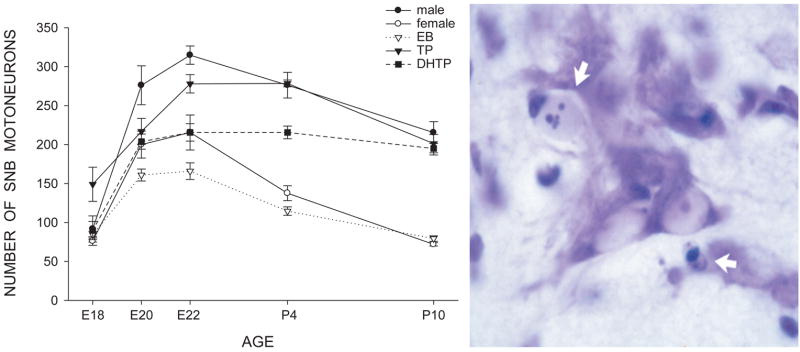
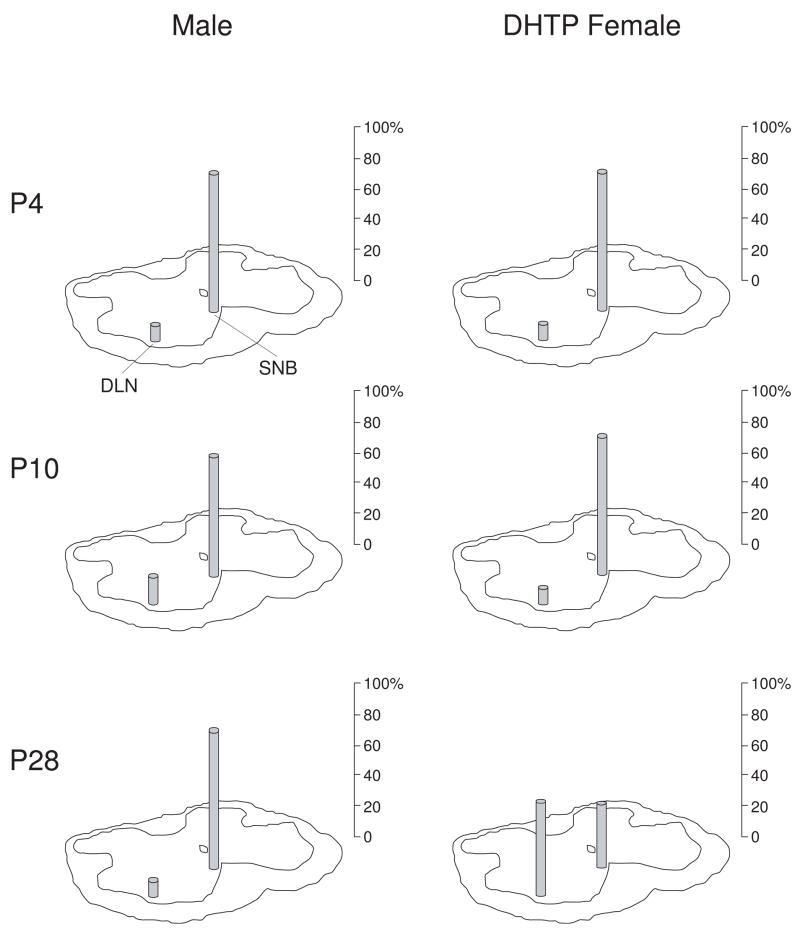
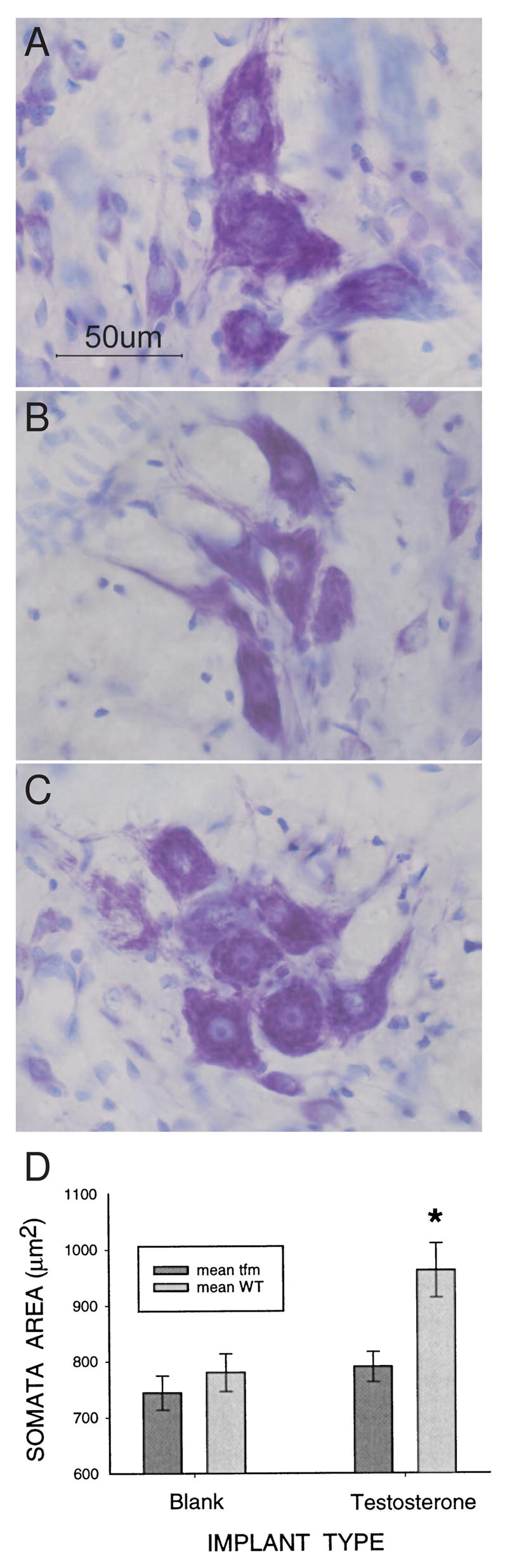
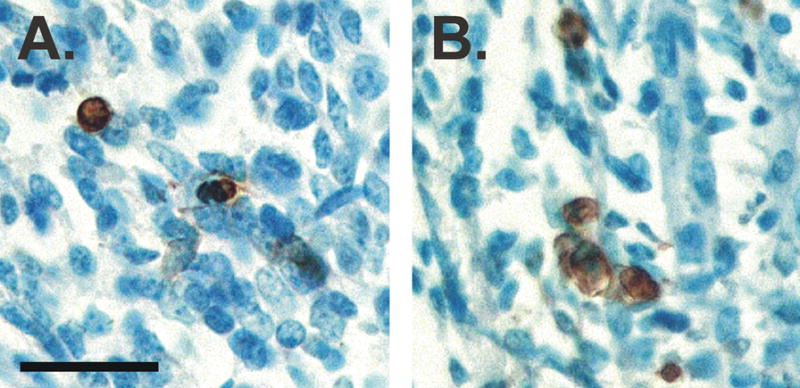
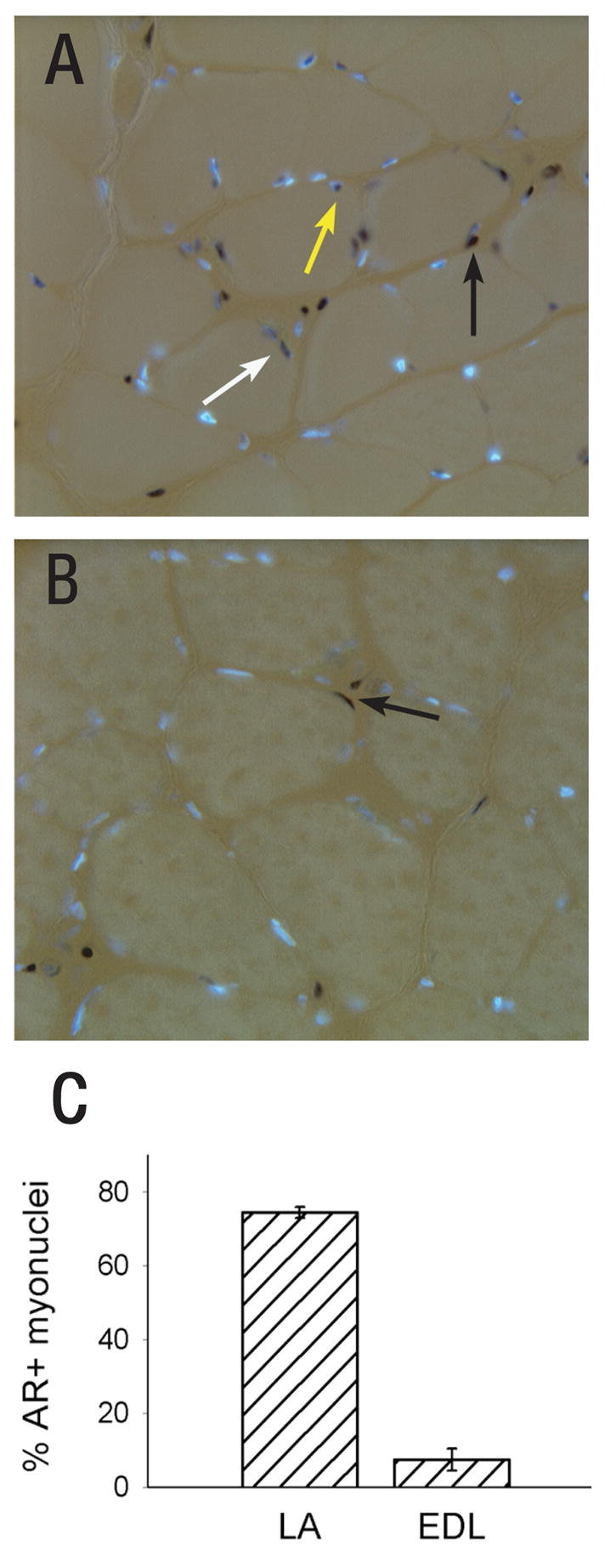
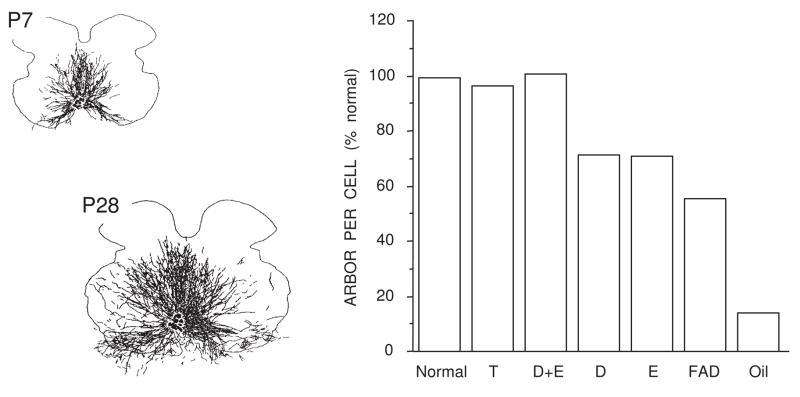
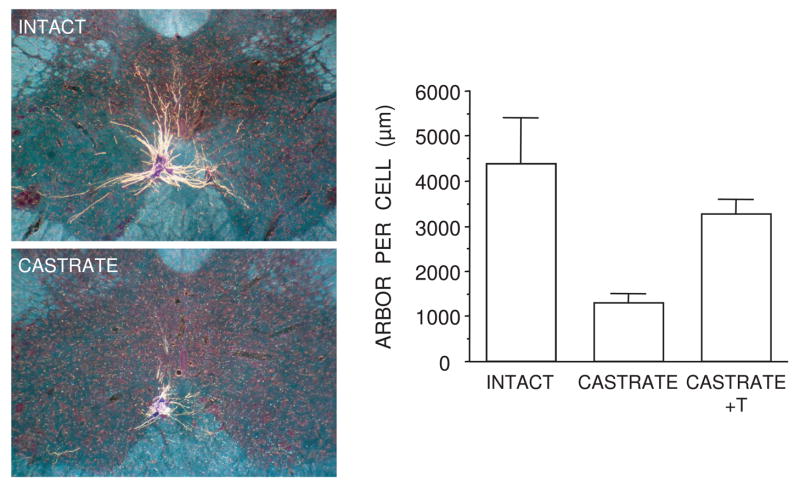
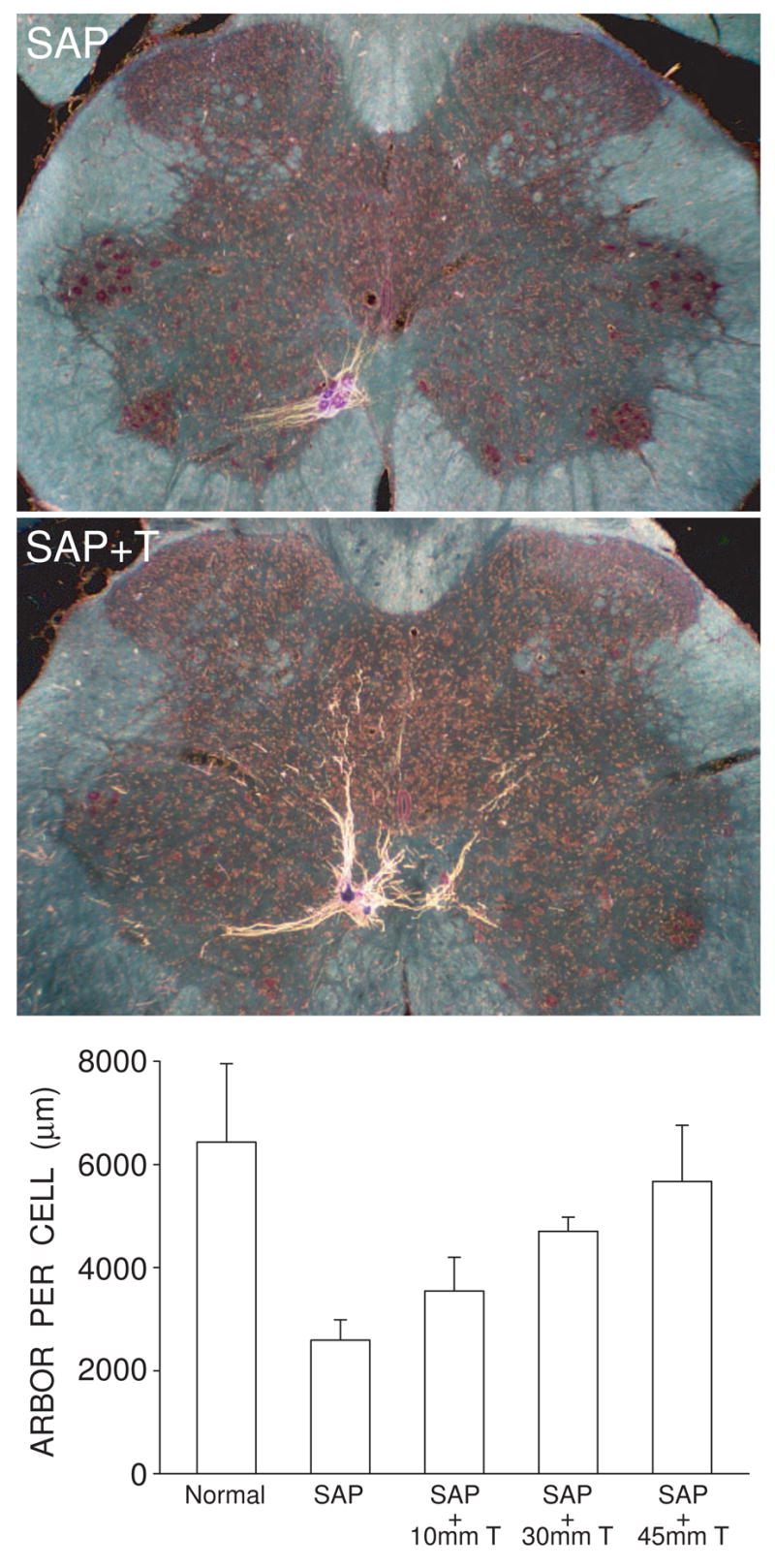
Cell number in the spinal nucleus of the bulbocavernosus (SNB) of rats was the first neural sex difference shown to differentiate under the control of androgens, acting via classical intracellular androgen receptors. SNB motoneurons reside in the lumbar spinal cord and innervate striated muscles involved in copulation, including the bulbocavernosus (BC) and levator ani (LA). SNB cells are much larger and more numerous in males than in females, and the BC/LA target muscles are reduced or absent in females. The relative simplicity of this neuromuscular system has allowed for considerable progress in pinpointing sites of hormone action, and identifying the cellular bases for androgenic effects. It is now clear that androgens act at virtually every level of the SNB system, in development and throughout adult life. In this review we focus on effects of androgens on developmental cell death of SNB motoneurons and BC/LA muscles; the establishment and maintenance of SNB motoneuron soma size and dendritic length; BC/LA muscle morphology and physiology; and behaviors controlled by the SNB system. We also describe new data on neurotherapeutic effects of androgens on SNB motoneurons after injury in adulthood.
Figures
Similar articles
-
Motoneuron morphology in the dorsolateral nucleus of the rat spinal cord: normal development and androgenic regulation.J Comp Neurol. 1993 Dec 22;338(4):588-600. doi: 10.1002/cne.903380408. J Comp Neurol. 1993. PMID: 7510730
-
Morphology of rat spinal motoneurons with normal and hormonally altered specificity.J Comp Neurol. 1990 Feb 22;292(4):638-50. doi: 10.1002/cne.902920412. J Comp Neurol. 1990. PMID: 2324318
-
Cellular analyses of hormone influence on motoneuronal development and function.J Neurobiol. 1986 May;17(3):157-76. doi: 10.1002/neu.480170304. J Neurobiol. 1986. PMID: 3519862 Review.
-
Characterization of the spinal nucleus of the bulbocavernosus neuromuscular system in male mice lacking androgen receptor in the nervous system.Endocrinology. 2012 Jul;153(7):3376-85. doi: 10.1210/en.2012-1001. Epub 2012 May 14. Endocrinology. 2012. PMID: 22585832
-
Regulation of motoneuron death in the spinal nucleus of the bulbocavernosus.J Neurobiol. 1992 Nov;23(9):1192-203. doi: 10.1002/neu.480230910. J Neurobiol. 1992. PMID: 1469384 Review.
Cited by
-
Turning sex inside-out: Peripheral contributions to sexual differentiation of the central nervous system.Biol Sex Differ. 2012 May 28;3(1):12. doi: 10.1186/2042-6410-3-12. Biol Sex Differ. 2012. PMID: 22640590 Free PMC article.
-
Patterns of connexin36 and eGFP reporter expression among motoneurons in spinal sexually dimorphic motor nuclei in mouse.Int J Physiol Pathophysiol Pharmacol. 2024 Jun 15;16(3):55-76. doi: 10.62347/OGWV9376. eCollection 2024. Int J Physiol Pathophysiol Pharmacol. 2024. PMID: 39021417 Free PMC article.
-
Sex differences in the brain: a whole body perspective.Biol Sex Differ. 2015 Aug 15;6:15. doi: 10.1186/s13293-015-0032-z. eCollection 2015. Biol Sex Differ. 2015. PMID: 26279833 Free PMC article. Review.
-
Sex Steroids as Regulators of Gestural Communication.Endocrinology. 2020 Jul 1;161(7):bqaa064. doi: 10.1210/endocr/bqaa064. Endocrinology. 2020. PMID: 32307535 Free PMC article. Review.
-
Sexually dimorphic nuclei in the spinal cord control male sexual functions.Front Neurosci. 2014 Jul 11;8:184. doi: 10.3389/fnins.2014.00184. eCollection 2014. Front Neurosci. 2014. PMID: 25071429 Free PMC article. Review.
References
-
- Bengston L, Lopez V, Watamura S, Forger NG. Short- and long-term effects of ciliary neurotrophic factor on androgen-sensitive motoneurons in the lumbar spinal cord. J Neurobiol. 1996;31:263–273. - PubMed
-
- Bleisch WV, Harrelson A. Androgens modulate endplate size and ACh receptor density at synapses in rat levator ani muscle. J Neurobiol. 1989;20:189–202. - PubMed
-
- Bleisch WV, Harrelson AL, Luine VN. Testosterone increases acetylcholine receptor number in the “levator ani” muscle of the rat. J Neurobiol. 1982;13:153–161. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
