

Murine beta-defensin 2 promotes TLR-4/MyD88-mediated and NF-kappaB-dependent atypical death of APCs via activation of TNFR2
- PMID: 18192488
- PMCID: PMC2365917
- DOI: 10.1189/jlb.1007700
Murine beta-defensin 2 promotes TLR-4/MyD88-mediated and NF-kappaB-dependent atypical death of APCs via activation of TNFR2
Abstract
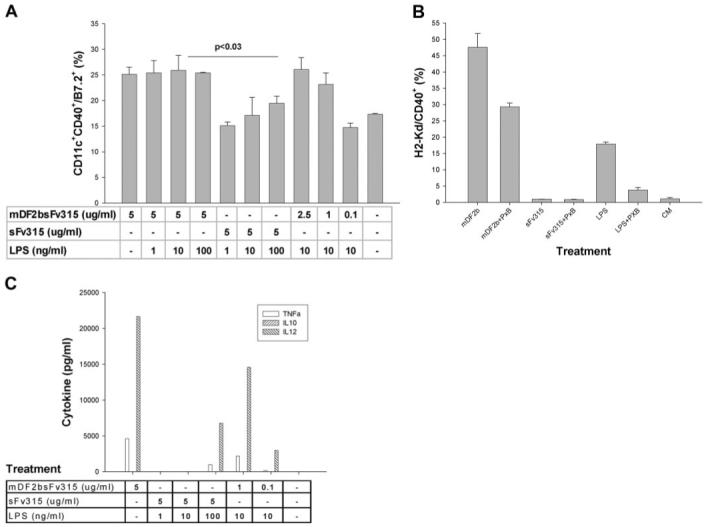
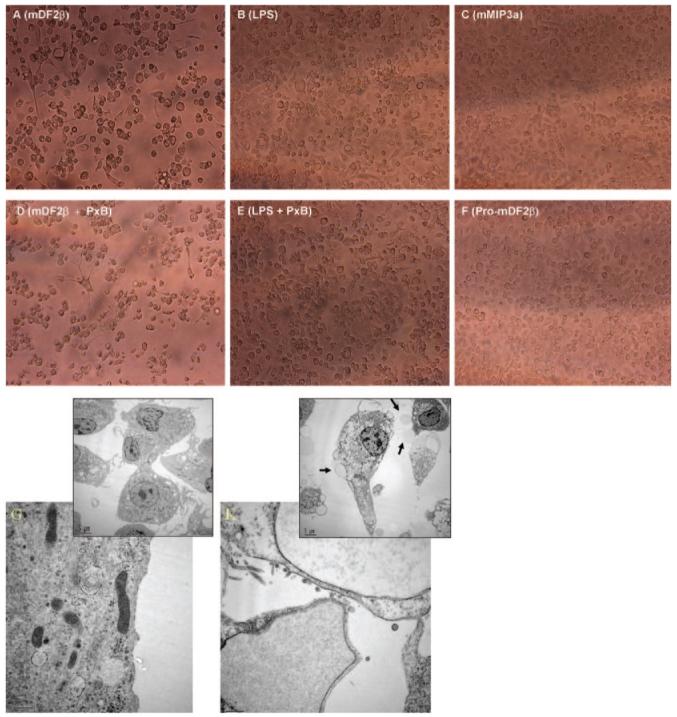
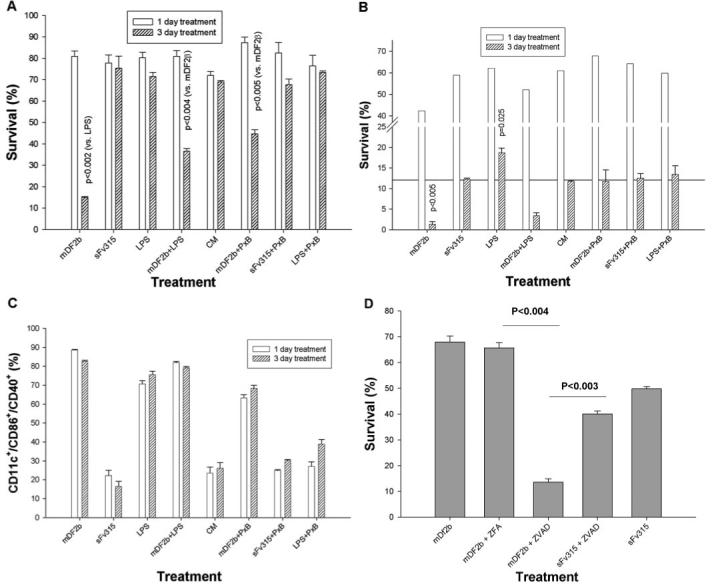
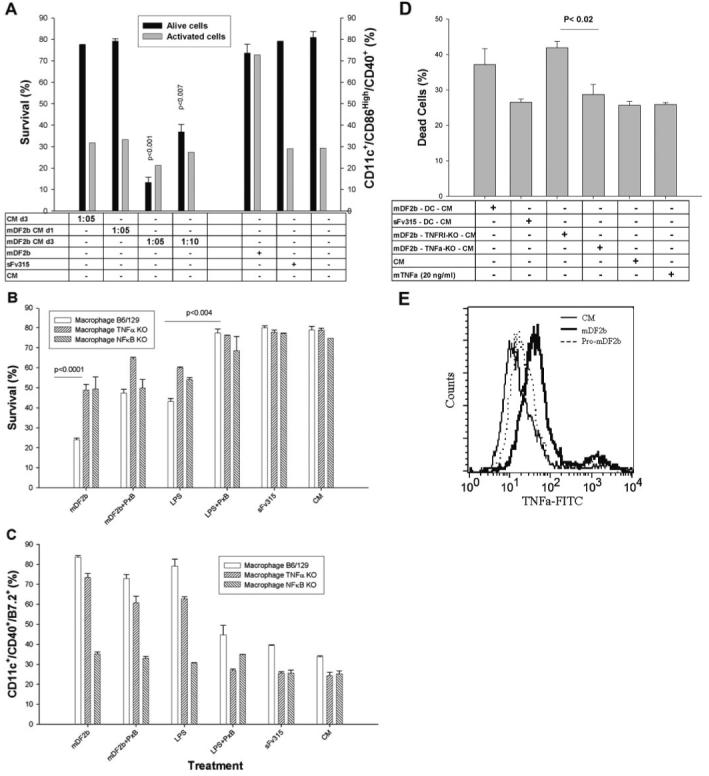
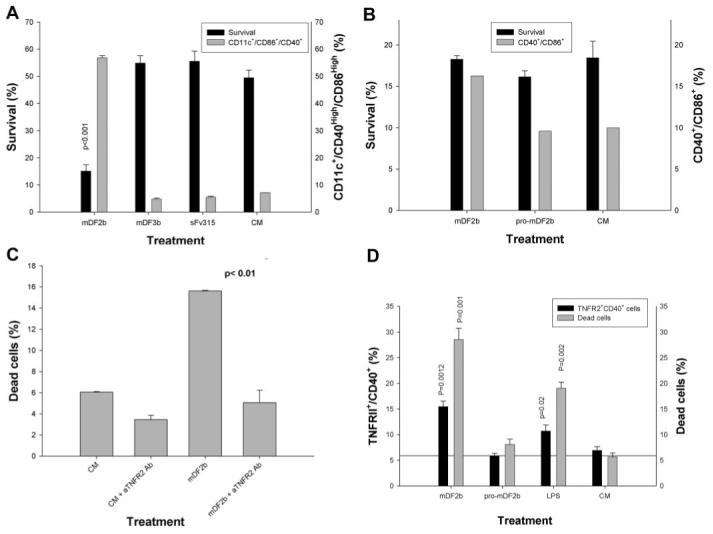
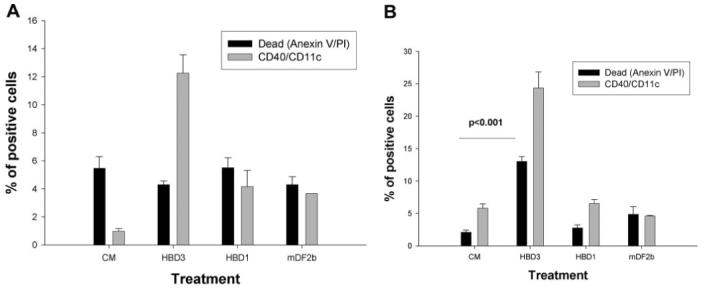
Mammalian antimicrobial peptides, including beta-defensins, represent an ancient arm of innate immunity designed to directly neutralize invading microbes. Previously, we demonstrated that murine beta-defensin 2 (mDF2beta) also acted as an endogenous ligand for TLR-4-activating maturation of dendritic cells (DCs). Herein, we report that this TLR-4 -dependent activation leads to induction of an atypical cell death that is unexpectedly exaggerated by the inhibition of caspases. Experiments using APCs with nonfunctional TNF-alpha or its receptors suggest that this is a NF-kappaB- and TNF-alpha-dependent process that does not require TNFR1. We demonstrate that mDF2beta triggers a TNFR2-mediated signaling cascade of "self-destruction" through up-regulation of membrane-bound TNF-alpha and TNFR2. This appears not to be an isolated phenomenon, as human synthetic beta-defenisn 3 was also able to activate and kill DCs. We propose that beta-defenins may play an important immunoregulatory role as controllers of the natural process of elimination of activated APCs.
Figures
Similar articles
-
Dual stimulation of MyD88-dependent Toll-like receptors induces synergistically enhanced production of inflammatory cytokines in murine bone marrow-derived dendritic cells.J Infect Dis. 2010 Jul 15;202(2):318-29. doi: 10.1086/653499. J Infect Dis. 2010. PMID: 20524851
-
Funiculosin variants and phosphorylated derivatives promote innate immune responses via the Toll-like receptor 4/myeloid differentiation factor-2 complex.J Biol Chem. 2017 Sep 15;292(37):15378-15394. doi: 10.1074/jbc.M117.791780. Epub 2017 Jul 28. J Biol Chem. 2017. PMID: 28754693 Free PMC article.
-
Synergistic BM-DC activation and immune induction by the oral vaccine vector Streptococcus gordonii and exogenous tumor necrosis factor.Mol Immunol. 2009 May;46(8-9):1883-91. doi: 10.1016/j.molimm.2009.02.008. Epub 2009 Mar 10. Mol Immunol. 2009. PMID: 19278729
-
Genetic and Epigenetic Regulation of the Innate Immune Response to Gout.Immunol Invest. 2023 Apr;52(3):364-397. doi: 10.1080/08820139.2023.2168554. Epub 2023 Feb 6. Immunol Invest. 2023. PMID: 36745138 Review.
-
Regulation of the MIR155 host gene in physiological and pathological processes.Gene. 2013 Dec 10;532(1):1-12. doi: 10.1016/j.gene.2012.12.009. Epub 2012 Dec 14. Gene. 2013. PMID: 23246696 Review.
Cited by
-
Novel apidaecin 1b analogs with superior serum stabilities for treatment of infections by gram-negative pathogens.Antimicrob Agents Chemother. 2013 Jan;57(1):402-9. doi: 10.1128/AAC.01923-12. Epub 2012 Oct 31. Antimicrob Agents Chemother. 2013. PMID: 23114765 Free PMC article.
-
Andrographolide mitigates IL‑1β‑induced human nucleus pulposus cells degeneration through the TLR4/MyD88/NF‑κB signaling pathway.Mol Med Rep. 2018 Dec;18(6):5427-5436. doi: 10.3892/mmr.2018.9599. Epub 2018 Oct 25. Mol Med Rep. 2018. PMID: 30365119 Free PMC article.
-
Strain-specific polymorphisms in Paneth cell α-defensins of C57BL/6 mice and evidence of vestigial myeloid α-defensin pseudogenes.Infect Immun. 2011 Jan;79(1):459-73. doi: 10.1128/IAI.00996-10. Epub 2010 Nov 1. Infect Immun. 2011. PMID: 21041494 Free PMC article.
-
Mechanisms and regulation of defensins in host defense.Signal Transduct Target Ther. 2023 Aug 14;8(1):300. doi: 10.1038/s41392-023-01553-x. Signal Transduct Target Ther. 2023. PMID: 37574471 Free PMC article. Review.
-
Endogenous toll-like receptor ligands and their biological significance.J Cell Mol Med. 2010 Nov;14(11):2592-603. doi: 10.1111/j.1582-4934.2010.01127.x. J Cell Mol Med. 2010. PMID: 20629986 Free PMC article. Review.
References
-
- Biragyn A. Defensins—non-antibiotic use for vaccine development. Curr. Protein Pept. Sci. 2005;6:53–60. - PubMed
-
- Ganz T. Defensins: antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003;3:710–720. - PubMed
-
- Yang D, Biragyn A, Hoover DM, Lubkowski J, Oppenheim JJ. Multiple roles of antimicrobial defensins, cathelicidins, and eosinophil-derived neurotoxin in host defense. Annu. Rev. Immunol. 2004;22:181–215. - PubMed
-
- Chaly YV, Paleolog EM, Kolesnikova TS, Tikhonov II, Petratchenko EV, Voitenok NN. Neutrophil α-defensin human neutrophil peptide modulates cytokine production in human monocytes and adhesion molecule expression in endothelial cells. Eur. Cytokine Netw. 2000;11:257–266. - PubMed
-
- van Wetering S, Sterk PJ, Rabe KF, Hiemstra PS. Defensins: key players or bystanders in infection, injury, and repair in the lung? J. Allergy Clin. Immunol. 1999;104:1131–1138. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
