

Review
doi: 10.1111/j.1600-0854.2008.00703.x.
Epub 2008 Jan 9.
Migration of Apicomplexa across biological barriers: the Toxoplasma and Plasmodium rides
Affiliations
- PMID: 18194412
- PMCID: PMC2329915
- DOI: 10.1111/j.1600-0854.2008.00703.x
Item in Clipboard
Review
Migration of Apicomplexa across biological barriers: the Toxoplasma and Plasmodium rides
Traffic.
2008 May.
Abstract
The invasive stages of Apicomplexa parasites, called zoites, have been largely studied in in vitro systems, with a special emphasis on their unique gliding and host cell invasive capacities. In contrast, the means by which these parasites reach their destination in their hosts are still poorly understood. We summarize here our current understanding of the cellular basis of in vivo parasitism by two well-studied Apicomplexa zoites, the Toxoplasma tachyzoite and the Plasmodium sporozoite. Despite being close relatives, these two zoites use different strategies to reach their goal and establish infection.
Figures
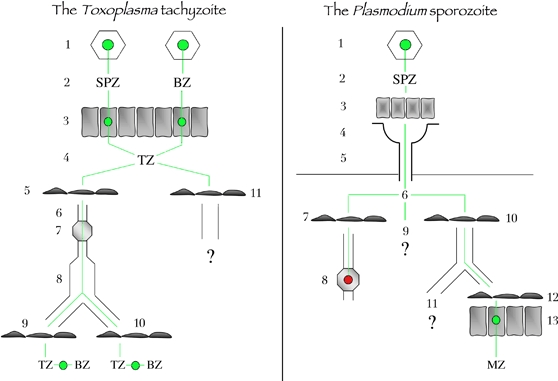
Left, the Toxoplasma tachyzoite. Toxoplasma infection starts by the ingestion of oocysts or tissues cysts (1) that liberate in the intestinal lumen free sporozoites or bradyzoites, respectively (2). Free parasites invade enterocytes (3), where they multiply and transform into tachyzoites, which are released in the lamina propria of the intestine (4). Tachyzoites invade the endothelium of lymphatic vessels (5), are taken up by the lymph (6), go through lymph nodes (7) and reach the blood circulation (8). Tachyzoites then cross blood endothelium barriers in immunoprivileged organs (9), for example, the brain, the retina or the placenta, or in other organs (10), for example, muscles, where they invade host cells, multiply and transform into dormant bradyzoites (11). Tachyzoites in the intestine may also cross the endothelium of blood vessels. Right, the Plasmodium sporozoite. Sporozoites are formed inside oocysts (1) in the wall of the mosquito midgut, are released in the hemocele bathed by the hemolymph (2), invade acinar cells (3) and exit in the secretory cavities (4) of the salivary glands and finally move into the secretory ducts (5). In the mammalian host, sporozoites are deposited in the dermis (6). Sporozoites can invade the endothelium of lymphatic vessels in the dermis (7) and then end up in the proximal draining lymph node (8), where they are killed. Sporozoites can remain in the dermis (9), in which case their fate is unknown. They can invade the endothelium of blood vessels in the dermis (10), reach the liver, cross the endothelium barrier of liver sinusoids (12) and invade hepatocytes (13), where they transform into merozoites, the erythrocyte-infecting form of the parasite. Sporozoites in the blood can also reach other organs than the liver (11), for example, the spleen, where the sporozoite fate is not well documented. BZ, bradyzoite; MZ, merozoite; SPZ, sporozoite; TZ, tachyzoite.
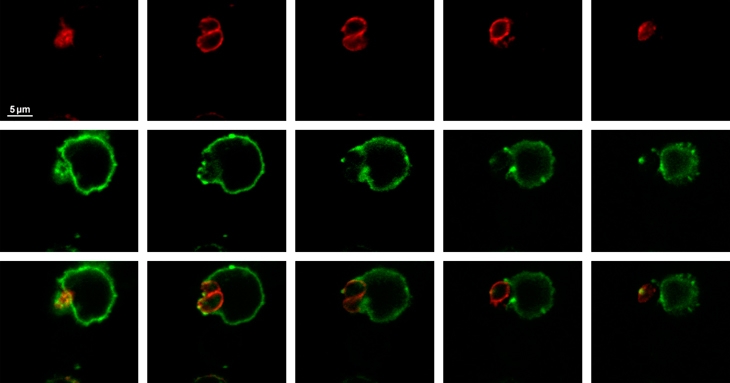
A mouse was infected with cysts of the 76K Toxoplasma gondii strain, mesenteric lymph nodes were recovered 5 days after infection, a cell suspension from the lymph node was fixed, MHC class II molecules were detected using class II I-Abdq and I-Edk antibodies (green), the parasite surface was stained using anti-SAG1 antibodies (red) and samples were observed by confocal microscopy (0.4-μm section). Note that the tachyzoites are located at the periphery of the host cell and are surrounded by the plasma membrane of the host cell. MHC, major histocompatibility complex.
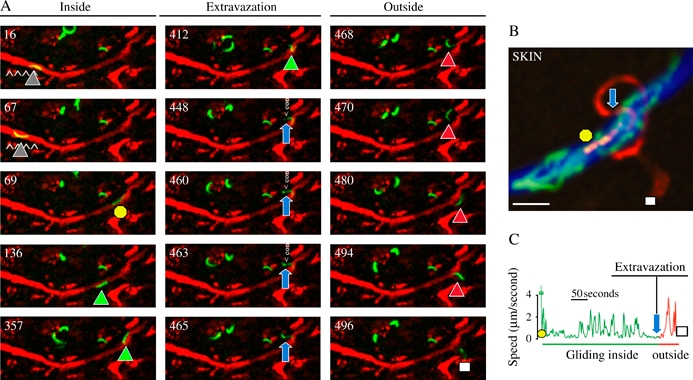
A) The fluorescent sporozoite glides inside the vessel colored in red after injection of red fluorescent BSA. The time (in seconds) is indicated in the upper left part of each panel. The intravascular sporozoite glides during the first 67 seconds (gray triangles), is suddenly displaced (yellow circle, 69 seconds), glides again inside the vessel (green triangles), extravazates (from 448 to 465 seconds, note the sporozoite constriction pointed by the blue arrows) before gliding in the dermis (468 to 496 seconds, red triangles until the white square). B) Maximum intensity projection of the fluorescent sporozoite from the 69th (yellow circle) to the 496th second (white square) through the constriction (blue arrow). C) Velocity profile of the sporozoite between the 69th and the 496th second.
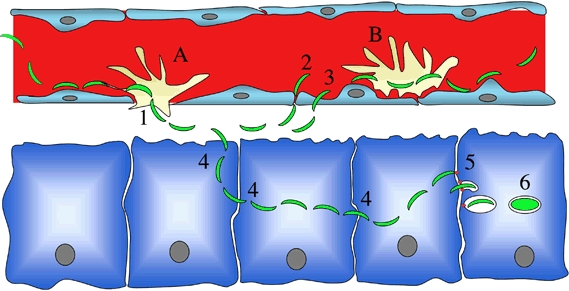
It is still unclear whether Kupffer cells are embedded in the sinusoidal barrier (A) or sit on the top of endothelial cells (B). If (A) occurs, then traversal of Kupffer cells (either by transcytosis, i.e., involving sporozoite entry inside a PV followed by escape from the cell, or by disrupting the Kupffer cell membranes and migrating through the cell) would be sufficient for crossing the sinusoidal barrier. If (B) occurs, then the sporozoite may, after traversing Kupffer cells, cross the endothelial barrier either by a paracellular route (2) or by traversing endothelial cells (3). Once in the parenchyma, the sporozoite traverses several hepatocytes (4) before invading a final one inside a PV (5), the only niche where a sporozoite can fully develop (6). In two opposing views, the final invasion step is either activated by prior hepatocyte traversal or instead constitutively available and requires inhibition of cell traversal.
References
-
- Dubremetz JF, Garcia-Réguet N, Conseil V, Fourmaux MN. Apical organelles and host cell invasion by Apicomplexa. Int J Parasitol. 1998;28:1007–1013. - PubMed
-
- Heintzelman MB. Cellular and molecular mechanisms of gliding locomotion in eukaryotes. Int Rev Cytol. 2006;251:79–129. - PubMed
-
- Keeley A, Soldati D. The glideosome: a molecular machine powering motility and host-cell invasion by Apicomplexa. Trends Cell Biol. 2004;14:528–532. - PubMed
-
- Fowler RE, Margos G, Mitchell GH. The cytoskeleton and motility in apicomplexan invasion. Adv Parasitol. 2004;56:213–263. - PubMed
-
- Mota MM, Rodriguez A. Migration through host cells by apicomplexan parasites. Microbes Infect. 2001;3:1123–1128. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
