

Review
doi: 10.1016/j.devcel.2007.12.008.
Viral and developmental cell fusion mechanisms: conservation and divergence
Affiliations
- PMID: 18194649
- PMCID: PMC3549671
- DOI: 10.1016/j.devcel.2007.12.008
Item in Clipboard
Review
Viral and developmental cell fusion mechanisms: conservation and divergence
Dev Cell.
2008 Jan.
Abstract
Membrane fusion is a fundamental requirement in numerous developmental, physiological, and pathological processes in eukaryotes. So far, only a limited number of viral and cellular fusogens, proteins that fuse membranes, have been isolated and characterized. Despite the diversity in structures and functions of known fusogens, some common principles of action apply to all fusion reactions. These can serve as guidelines in the search for new fusogens, and may allow the formulation of a cross-species, unified theory to explain divergent and convergent evolutionary principles of membrane fusion.
Figures
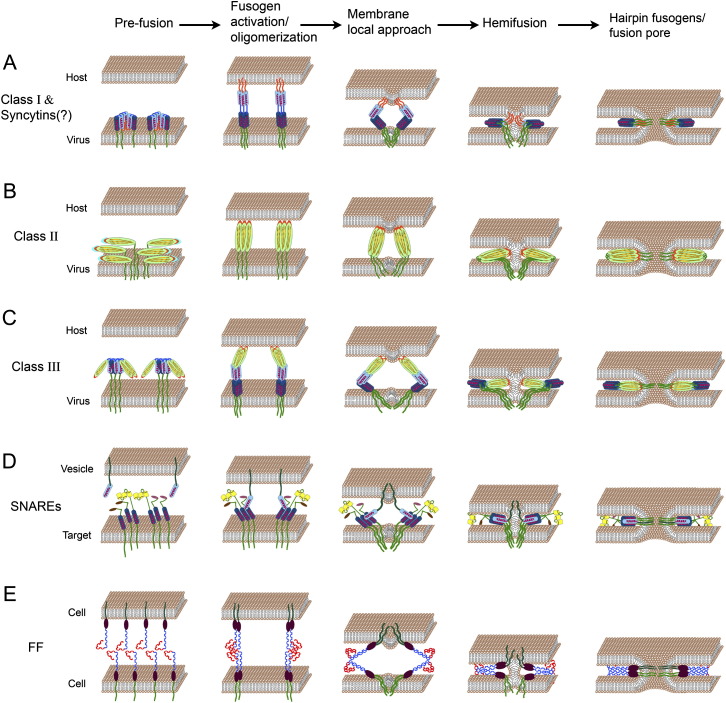
Models for Membrane Fusion Mediated by Viral, Intracellular, and Developmental Fusogens Fusogens are drawn embedded in the membranes after the stages of oligomerization, sorting, trafficking, and binding to the host receptors (not shown). α helices, light and dark blue cylinders; transmembrane domains, dark green; fusion peptides or loops, orange; β sheets, light green ellipses. (A) Class I viral fusogens (such as in HIV) contain two α helices and an amphiphilic fusion peptide buried inside the protein in the prefusion state. Upon activation, the trimers undergo a conformational change leading to the formation of rigid coiled-coil structures with exposed fusion peptides at their N termini. Insertion of fusion peptides into the host membrane, concomitant with protein fold back into six-helix bundles, leads to membrane tightening, hemifusion, and pore formation (Kielian and Rey, 2006). (B) In Class II viral fusogens (for example, in Dengue viruses), hydrophobic fusion loops emanate from a structure containing β sheets. After fusogen activation and a shift from heterodimers to homotrimers (not shown), the fusion loops are exposed and inserted into the opposing membrane. Folding back occurs presumably by interaction between different domains of the protein that drive membrane fusion (Kielian and Rey, 2006). (C) Class III fusogens (such as in Rabies and VSV viruses) combine characteristics of Class I and II including short and discontinuous hydrophobic fusion loops that stem from a structure containing β sheets, and α helices that potentially form six-helix bundles to merge the membranes together. In contrast to class I and II fusogens, activation does not require fusion domain exposure (Heldwein et al., 2006, Weissenhorn et al., 2007). (D) SNARE-dependent intracellular fusion. Three different t-SNAREs, each with a single α helix domain, are bundled to form the target complex. This complex binds a single α helix of v-SNARE that emanates from the vesicle membrane. Transition between loose and tight t- SNARE-v-SNARE complexes is mediated by SNARE zippering and tethering of the membranes, leading to their fusion (Jahn and Scheller, 2006). (E) Hypothetical model of FF fusion. The extracellular domains of FF proteins (C. elegans EFF-1 and AFF-1 proteins) are comprised of juxta-membrane hypothetical globular domains (brown ellipses) and a putative cysteine-rich conserved domain that presumably forms an S-S based structure (in blue). Part of these predicted extracellular loops are reminiscent of TGF-β receptor-like domains (in red). We hypothesize that cis-dimerization of FF molecules is followed by homotypic trans-binding between dimmers. Trans-zippering of FF complexes from the two membranes induces membrane tightening, hemifusion, and fusion pore formation.
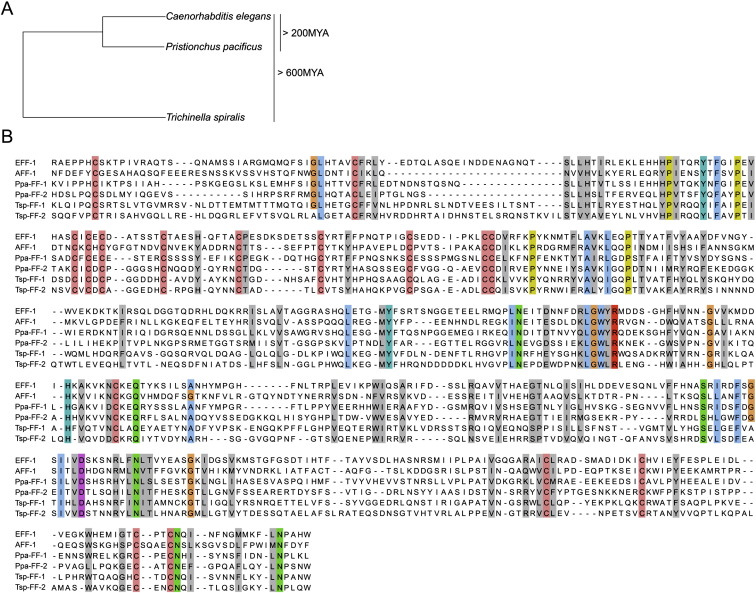
Conservation of Residues and Motifs in FF Proteins from Nematodes (A) Phylogeny of nematode species used for the sequence alignment. Using BLAST, FF proteins were identified in 17 nematode species from different clades (O.A. and B.P., unpublished data). We aligned Caenorhabditis elegans (Ce) FF proteins with putative homologs from Pristionchus pacificus (Ppa), estimated to diverge 200 million years ago (MYA) (Gutierrez and Sommer, 2004), and with proteins from the remotest nematode wherein FF proteins have been identified so far, the parasite Trichinella spiralis (Tsp) (divergence estimated more then 600 MYA; Mitreva and Jasmer, 2006). (B) Extracellular domains of FF proteins without signal sequences were aligned using the Jalview software (Clamp et al., 2004). Conservation of cysteines (pink), the LGWYR motifs, and partial conservation of prolines (yellow) suggests functional roles for these residues and motifs (Sapir et al., 2007). Color is shown in residues of 100% conservation and gray in cases where 50% of the physicochemical properties are conserved (Livingstone and Barton, 1993). Accession numbers: CeAFF-1: EF205023; CeEFF-1: C26D10.5 PpaFF-1: contig162.29; PpaFF-2: Contig735.1 P. pacificus california “Assembly Freeze 1.” TspFF-1: Contig1.317; TspFF-2: Contig3.96 Trichinella spiralis-1.0-contigs.
References
-
- Alper S., Kenyon C. REF-1, a protein with two bHLH domains, alters the pattern of cell fusion in C. elegans by regulating Hox protein activity. Development. 2001;128:1793–1804. - PubMed
-
- Alper S., Kenyon C. The zinc finger protein REF-2 functions with the Hox genes to inhibit cell fusion in the ventral epidermis of C. elegans. Development. 2002;129:3335–3348. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
