

A syntaxin 10-SNARE complex distinguishes two distinct transport routes from endosomes to the trans-Golgi in human cells
- PMID: 18195106
- PMCID: PMC2213607
- DOI: 10.1083/jcb.200707136
A syntaxin 10-SNARE complex distinguishes two distinct transport routes from endosomes to the trans-Golgi in human cells
Abstract
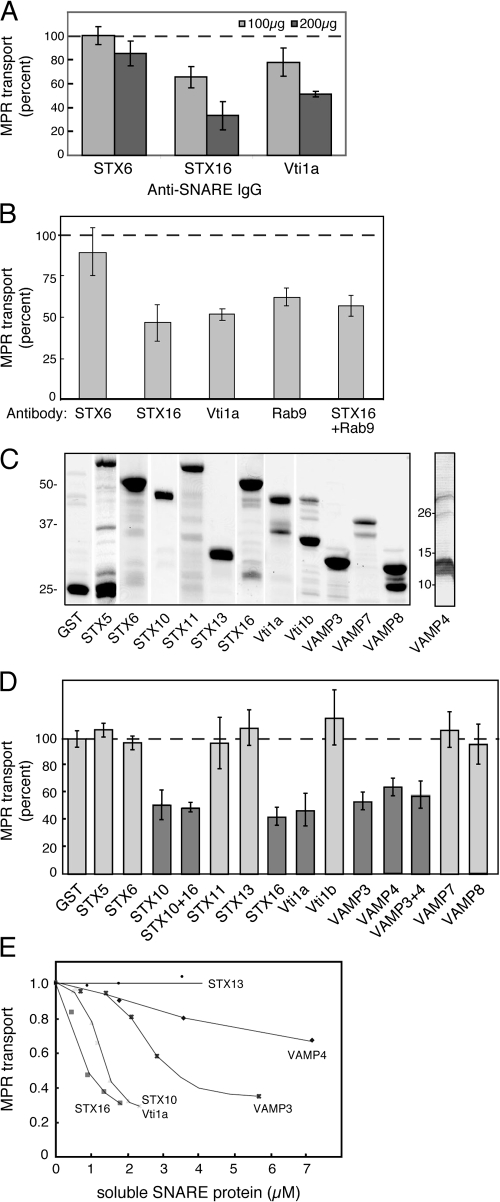
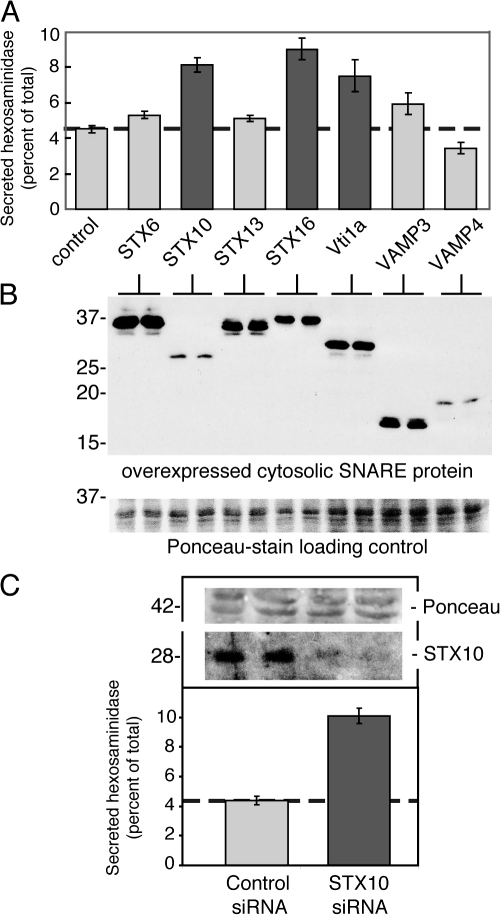
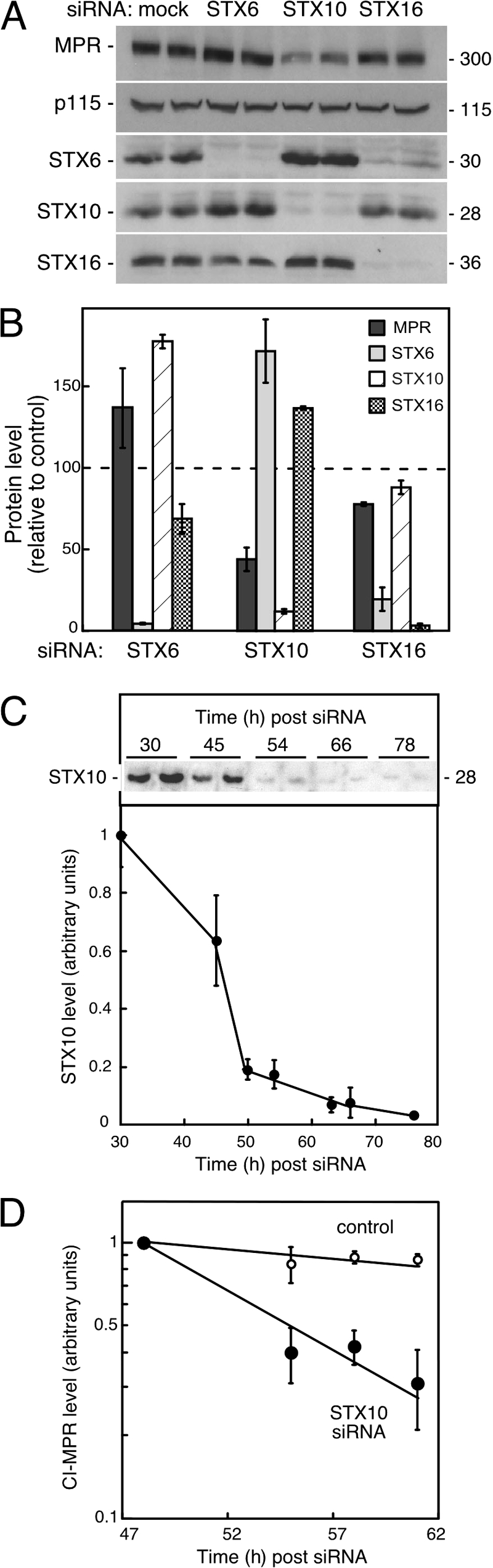
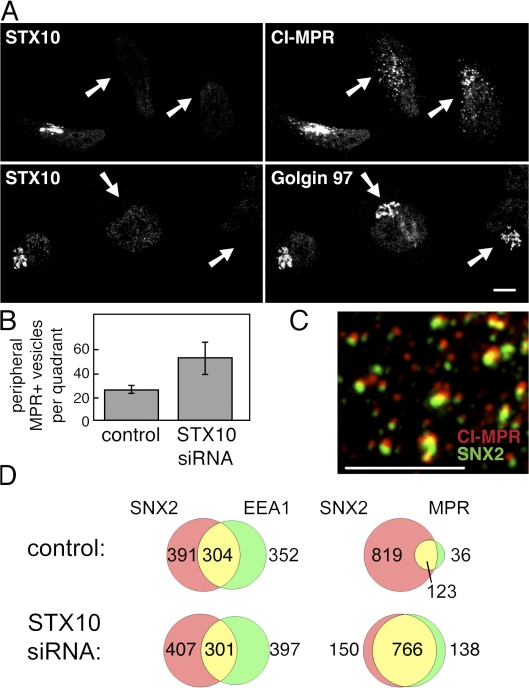
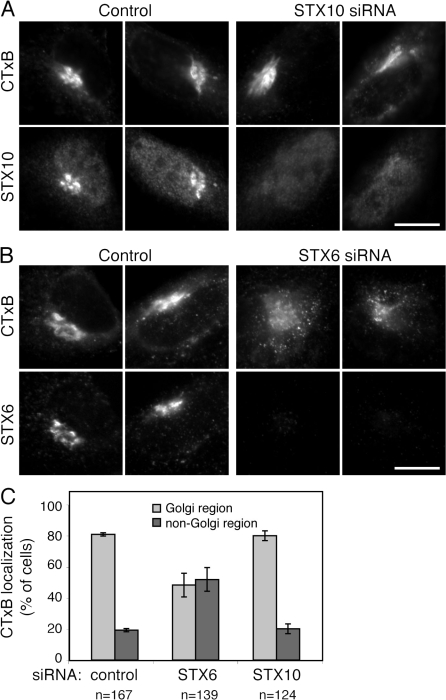
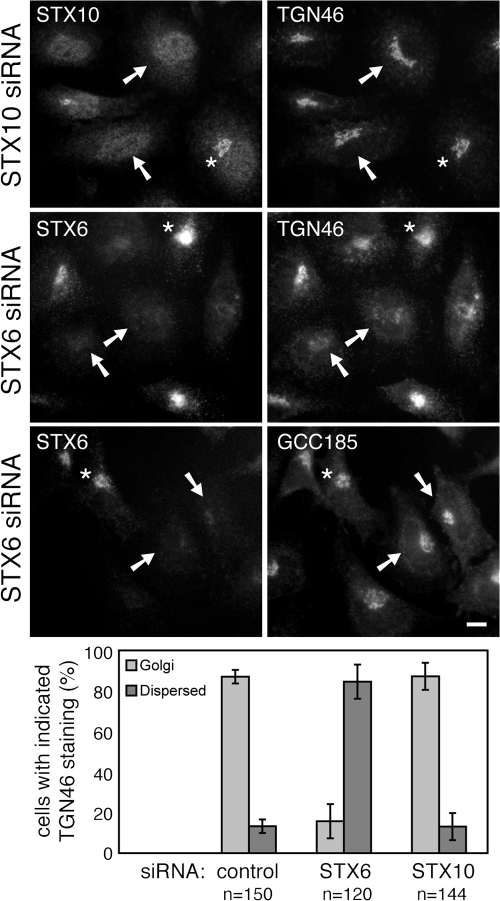
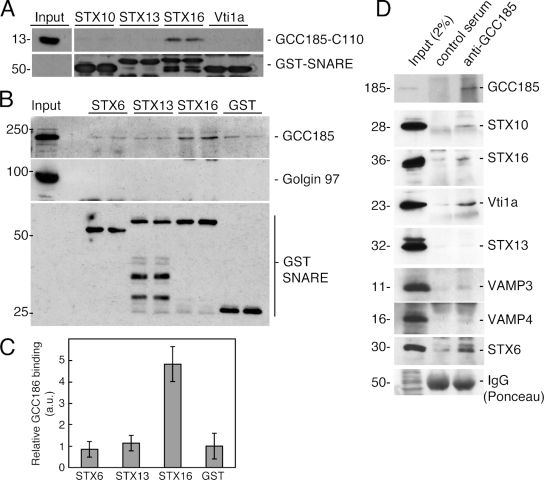
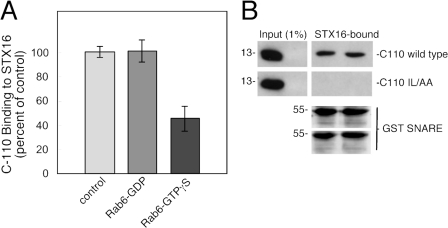
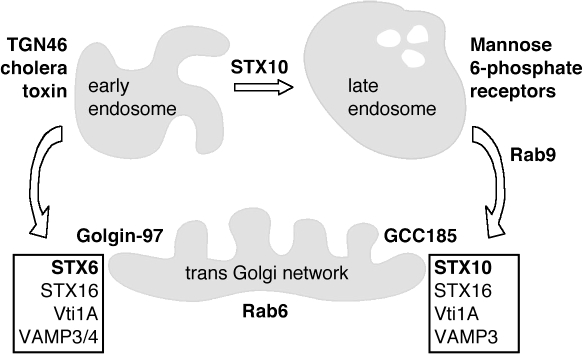
Mannose 6-phosphate receptors (MPRs) are transported from endosomes to the Golgi after delivering lysosomal enzymes to the endocytic pathway. This process requires Rab9 guanosine triphosphatase (GTPase) and the putative tether GCC185. We show in human cells that a soluble NSF attachment protein receptor (SNARE) complex comprised of syntaxin 10 (STX10), STX16, Vti1a, and VAMP3 is required for this MPR transport but not for the STX6-dependent transport of TGN46 or cholera toxin from early endosomes to the Golgi. Depletion of STX10 leads to MPR missorting and hypersecretion of hexosaminidase. Mouse and rat cells lack STX10 and, thus, must use a different target membrane SNARE for this process. GCC185 binds directly to STX16 and is competed by Rab6. These data support a model in which the GCC185 tether helps Rab9-bearing transport vesicles deliver their cargo to the trans-Golgi and suggest that Rab GTPases can regulate SNARE-tether interactions. Importantly, our data provide a clear molecular distinction between the transport of MPRs and TGN46 to the trans-Golgi.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
