

HSP90 regulates cell survival via inositol hexakisphosphate kinase-2
- PMID: 18195352
- PMCID: PMC2234104
- DOI: 10.1073/pnas.0711168105
HSP90 regulates cell survival via inositol hexakisphosphate kinase-2
Abstract
Heat-shock proteins (HSPs) are abundant, inducible proteins best known for their ability to maintain the conformation of proteins and to refold damaged proteins. Some HSPs, especially HSP90, can be antiapoptotic and the targets of anticancer drugs. Inositol hexakisphosphate kinase-2 (IP6K2), one of a family of enzymes generating the inositol pyrophosphate IP7 [diphosphoinositol pentakisphosphate (5-PP-IP5)], mediates apoptosis. Increased IP6K2 activity sensitizes cancer cells to stressors, whereas its depletion blocks cell death. We now show that HSP90 physiologically binds IP6K2 and inhibits its catalytic activity. Drugs and selective mutations that abolish HSP90-IP6K2 binding elicit activation of IP6K2, leading to cell death. Thus, the prosurvival actions of HSP90 reflect the inhibition of IP6K2, suggesting that selectively blocking this interaction could provide effective and safer modes of chemotherapy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
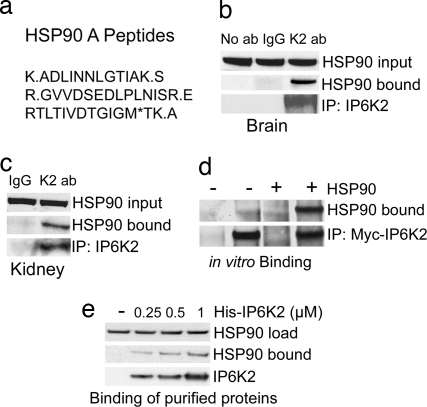
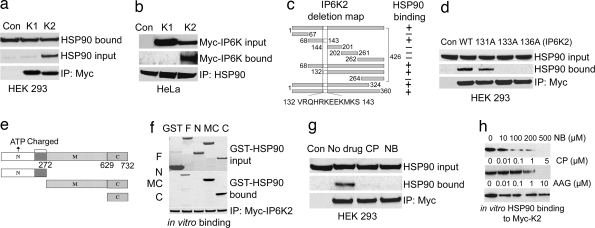
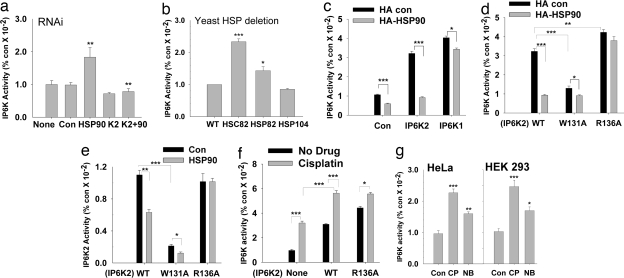
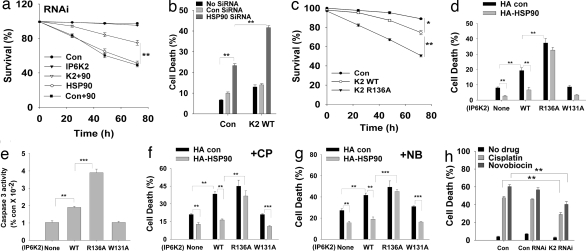
Comment in
-
IP6K2 is a client for HSP90 and a target for cancer therapeutics development.Proc Natl Acad Sci U S A. 2008 Feb 5;105(5):1389-90. doi: 10.1073/pnas.0711993105. Epub 2008 Jan 29. Proc Natl Acad Sci U S A. 2008. PMID: 18230718 Free PMC article. No abstract available.
References
-
- Berridge MJ, Lipp P, Bootman MD. Signal transduction: The calcium entry pas de deux. Science. 2000;287:1604–1605. - PubMed
-
- Irvine RF, Schell MJ. Back in the water: The return of the inositol phosphates. Nat Rev Mol Cell Biol. 2001;2:327–338. - PubMed
-
- Menniti FS, Miller RN, Putney JW, Jr, Shears SB. Turnover of inositol polyphosphate pyrophosphates in pancreatoma cells. J Biol Chem. 1993;268:3850–3856. - PubMed
-
- Stephens L, et al. The detection, purification, structural characterization, and metabolism of diphosphoinositol pentakisphosphate(s) and bisdiphosphoinositol tetrakisphosphate(s). J Biol Chem. 1993;268:4009–4015. - PubMed
-
- Saiardi A, Erdjument-Bromage H, Snowman AM, Tempst P, Snyder SH. Synthesis of diphosphoinositol pentakisphosphate by a newly identified family of higher inositol polyphosphate kinases. Curr Biol. 1999;9:1323–1326. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
