

Self-association of human PCSK9 correlates with its LDLR-degrading activity
- PMID: 18197702
- PMCID: PMC2732112
- DOI: 10.1021/bi7016359
Self-association of human PCSK9 correlates with its LDLR-degrading activity
Abstract
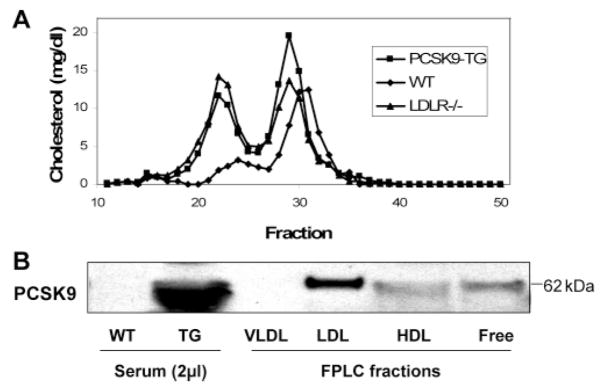
Genetic studies have demonstrated an important role for proprotein convertase subtilisin/kexin type 9 (PCSK9) as a determinant of plasma cholesterol levels. However, the underlying molecular mechanism is not completely understood. To this end, we have generated a mammalian cell expression system for human PCSK9 and its mutants and produced transgenic mice expressing human PCSK9. HEK293T cells transfected with the human PCSK9 DNA construct expressed and secreted PCSK9 and displayed decreased LDLR levels; functional PCSK9 protein was purified from the conditioned medium. In vitro studies showed that PCSK9 self-associated in a concentration-, temperature-, and pH-dependent manner. A mixture of PCSK9 monomers, dimers, and trimers displayed an enhanced LDLR degrading activity compared to monomeric PCSK9. A gain-of-function mutant, D374Y, displayed greatly increased self-association compared to wild-type PCSK9. Moreover, we demonstrated that the catalytic domain of PCSK9 is responsible for the self-association. Self-association of PCSK9 was enhanced by incubation with mouse apoE-/- VLDL and inhibited by incubation with both human and mouse HDL. When PCSK9 protein was incubated with total serum, it partially associated with LDL and HDL but not with VLDL. In transgenic mice, PCSK9 also associated with LDL and HDL but not with VLDL. We conclude that self-association is an intrinsic property of PCSK9, correlated to its LDLR-degrading activity and affected by plasma lipoproteins. These results provide a basis for developing strategies to manipulate PCSK9 activity in the circulation for the treatment of hypercholesterolemia.
Figures








References
-
- Naureckiene S, Ma L, Sreekumar K, Purandare U, Lo CF, Huang Y, Chiang LW, Grenier JM, Ozenberger BA, Jacobsen JS, Kennedy JD, DiStefano PS, Wood A, Bingham B. Functional characterization of Narc 1, a novel proteinase related to proteinase K. Arch Biochem Biophys. 2003;420:55–67. - PubMed
-
- Steiner D. The proprotein convertases. Curr Opin Chem Biol. 1998;2:31–39. - PubMed
-
- Zhou A, Webb G, Zhu X, Steiner DF. Proteolytic processing in the secretory pathway. J Biol Chem. 1999;274:20745–20748. - PubMed
-
- Seidah NG, Chretien M. Proprotein and prohormone convertases: a family of subtilases generating diverse bioactive polypeptides. Brain Res. 1999;848:45–62. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
