

T cell receptor microcluster transport through molecular mazes reveals mechanism of translocation
- PMID: 18199675
- PMCID: PMC2275686
- DOI: 10.1529/biophysj.107.119099
T cell receptor microcluster transport through molecular mazes reveals mechanism of translocation
Abstract
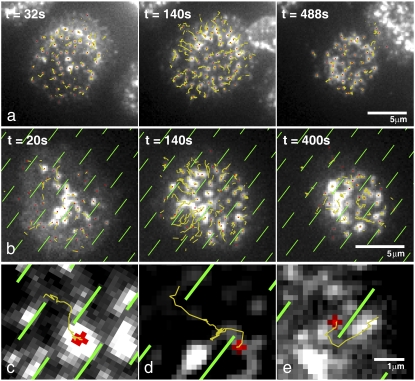
Recognition of peptide antigen by T cells involves coordinated movement of T cell receptors (TCRs) along with other costimulatory and signaling molecules. The spatially organized configurations that result are collectively referred to as the immunological synapse. Experimental investigation of the role of spatial organization in TCR signaling has been facilitated by the use of nanopatterned-supported membranes to direct TCR into alternative patterns. Here we study the mechanism by which substrate structures redirect TCR transport. Using a flow-tracking algorithm, the ensemble of TCR clusters within each cell was tracked during synapse formation under various constraint geometries. Shortly after initial cluster formation, a coordinated centripetal flow of approximately 20 nm/s develops. Clusters that encounter substrate-imposed constraint are deflected and move parallel to the constraint at speeds that scale with the relative angle of motion to the preferred centripetal direction. TCR transport is driven by actin polymerization, and the distribution of F-actin was imaged at various time points during the synapse formation process. At early time points, there is no significant effect on actin distribution produced by substrate constraints. At later time points, modest differences were observed. These data are consistent with a frictional model of TCR coupling to cytoskeletal flow, which allows slip. Implications of this model regarding spatial sorting of cell-surface molecules are discussed.
Figures





References
-
- Akins, M. R., and T. Biederer. 2006. Cell-cell interactions in synaptogenesis. Curr. Opin. Neurobiol. 16:83–89. - PubMed
-
- Krummel, M. F., and I. Macara. 2006. Maintenance and modulation of T cell polarity. Nat. Immunol. 7:1143–1149. - PubMed
-
- Grakoui, A., S. K. Bromley, C. Sumen, M. M. Davis, A. S. Shaw, P. M. Allen, and M. L. Dustin. 1999. The immunological synapse: A molecular machine controlling T cell activation. Science. 285:221–227. - PubMed
-
- Krummel, M. F., M. D. Sjaastad, C. Wulfing, and M. M. Davis. 2000. Differential clustering of CD4 and CD3 zeta during T cell recognition. Science. 289:1349–1352. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
