

Distinct chromatin modulators regulate the formation of accessible and repressive chromatin at the fission yeast recombination hotspot ade6-M26
- PMID: 18199689
- PMCID: PMC2262960
- DOI: 10.1091/mbc.e07-04-0377
Distinct chromatin modulators regulate the formation of accessible and repressive chromatin at the fission yeast recombination hotspot ade6-M26
Abstract
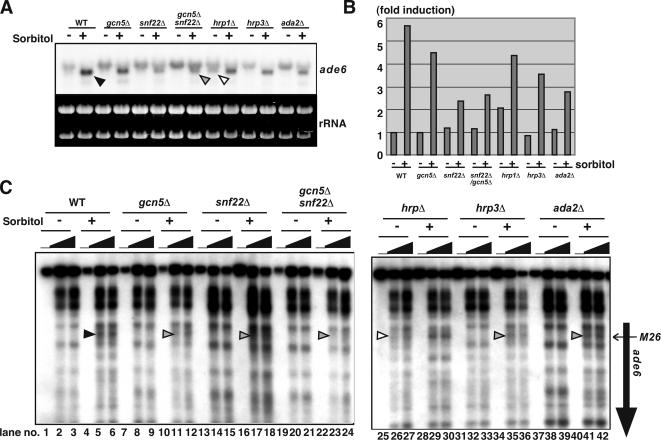
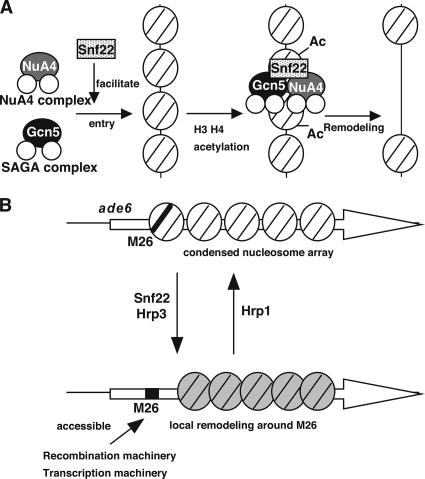
Histone acetyltransferases (HATs) and ATP-dependent chromatin remodeling factors (ADCRs) regulate transcription and recombination via alteration of local chromatin configuration. The ade6-M26 allele of Schizosaccharomyces pombe creates a meiotic recombination hotspot that requires a cAMP-responsive element (CRE)-like sequence M26, the Atf1/Pcr1 heterodimeric ATF/CREB transcription factor, the Gcn5 HAT, and the Snf22 SWI2/SNF2 family ADCR. Chromatin alteration occurs meiotically around M26, leading to the activation of meiotic recombination. We newly report the roles of other chromatin remodeling factors that function positively and negatively in chromatin alteration at M26: two CHD-1 family ADCRs (Hrp1 and Hrp3), a Spt-Ada-Gcn5 acetyltransferase component (Ada2), and a member of Moz-Ybf2/Sas3-Sas2-Tip60 family (Mst2). Ada2, Mst2, and Hrp3 are required for the full activation of chromatin changes around M26 and meiotic recombination. Acetylation of histone H3 around M26 is remarkably reduced in gcn5Delta, ada2Delta and snf22Delta, suggesting cooperative functions of these HAT complexes and Snf22. Conversely, Hrp1, another CHD-1 family ADCR, maintains repressive chromatin configuration at ade6-M26. Interestingly, transcriptional initiation site is shifted to a site around M26 from the original initiation sites, in couple with the histone acetylation and meiotic chromatin alteration induced around 3' region of M26, suggesting a collaboration between these chromatin modulators and the transcriptional machinery to form accessible chromatin. These HATs and ADCRs are also required for the regulation of transcription and chromatin structure around M26 in response to osmotic stress. Thus, we propose that multiple chromatin modulators regulate chromatin structure reversibly and participate in the regulation of both meiotic recombination and stress-induced transcription around CRE-like sequences.
Figures
Similar articles
-
Roles of histone acetylation and chromatin remodeling factor in a meiotic recombination hotspot.EMBO J. 2004 Apr 21;23(8):1792-803. doi: 10.1038/sj.emboj.7600138. Epub 2004 Feb 26. EMBO J. 2004. PMID: 14988732 Free PMC article.
-
Multiple modes of chromatin configuration at natural meiotic recombination hot spots in fission yeast.Eukaryot Cell. 2007 Nov;6(11):2072-80. doi: 10.1128/EC.00246-07. Epub 2007 Sep 7. Eukaryot Cell. 2007. PMID: 17827346 Free PMC article.
-
Interplay between chromatin modulators and histone acetylation regulates the formation of accessible chromatin in the upstream regulatory region of fission yeast fbp1.Genes Genet Syst. 2018 May 3;92(6):267-276. doi: 10.1266/ggs.17-00018. Epub 2017 Jun 30. Genes Genet Syst. 2018. PMID: 28674280
-
Regulation Mechanisms of Meiotic Recombination Revealed from the Analysis of a Fission Yeast Recombination Hotspot ade6-M26.Biomolecules. 2022 Nov 26;12(12):1761. doi: 10.3390/biom12121761. Biomolecules. 2022. PMID: 36551189 Free PMC article. Review.
-
The meiotic recombination hotspots of Schizosaccharomyces pombe.Genome Dyn. 2009;5:1-13. doi: 10.1159/000166614. Genome Dyn. 2009. PMID: 18948703 Review.
Cited by
-
New paradigms for conserved, multifactorial, cis-acting regulation of meiotic recombination.Nucleic Acids Res. 2012 Nov 1;40(20):9983-9. doi: 10.1093/nar/gks761. Epub 2012 Aug 16. Nucleic Acids Res. 2012. PMID: 22904082 Free PMC article. Review.
-
Molecular mechanisms for environmentally induced and evolutionarily rapid redistribution (plasticity) of meiotic recombination.Genetics. 2022 Feb 4;220(2):iyab212. doi: 10.1093/genetics/iyab212. Genetics. 2022. PMID: 34888655 Free PMC article.
-
Transcription of mRNA-type long non-coding RNAs (mlonRNAs) disrupts chromatin array.Commun Integr Biol. 2009;2(1):25-6. doi: 10.4161/cib.2.1.7378. Commun Integr Biol. 2009. PMID: 19704860 Free PMC article.
-
Anatomy of mouse recombination hot spots.Nucleic Acids Res. 2010 Apr;38(7):2346-54. doi: 10.1093/nar/gkp1251. Epub 2010 Jan 15. Nucleic Acids Res. 2010. PMID: 20081202 Free PMC article.
-
Adaptive Control of the Meiotic Recombination Landscape by DNA Site-dependent Hotspots With Implications for Evolution.Front Genet. 2022 Jun 22;13:947572. doi: 10.3389/fgene.2022.947572. eCollection 2022. Front Genet. 2022. PMID: 35812747 Free PMC article.
References
-
- Agalioti T., Lomvardas S., Parekh B., Yie J., Maniatis T., Thanos D. Ordered recruitment of chromatin modifying and general transcription factors to the IFN-beta promoter. Cell. 2000;103:667–678. - PubMed
-
- Bird A. W., Yu D. Y., Pray-Grant M. G., Qiu Q., Harmon K. E., Megee P. C., Grant P. A., Smith M. M., Christman M. F. Acetylation of histone H4 by Esa1 is required for DNA double-strand break repair. Nature. 2002;419:411–415. - PubMed
-
- Brown C. E., Lechner T., Howe L., Workman J. L. The many HATs of transcription coactivators. Trends Biochem. Sci. 2000;25:15–19. - PubMed
-
- Chen H., Tini M., Evans R. M. HATs on and beyond chromatin. Curr. Opin. Cell Biol. 2001;13:218–224. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
