

Regulation of respiratory and vocal motor pools in the isolated brain of Xenopus laevis
- PMID: 18199762
- PMCID: PMC2905839
- DOI: 10.1523/JNEUROSCI.4754-07.2008
Regulation of respiratory and vocal motor pools in the isolated brain of Xenopus laevis
Abstract
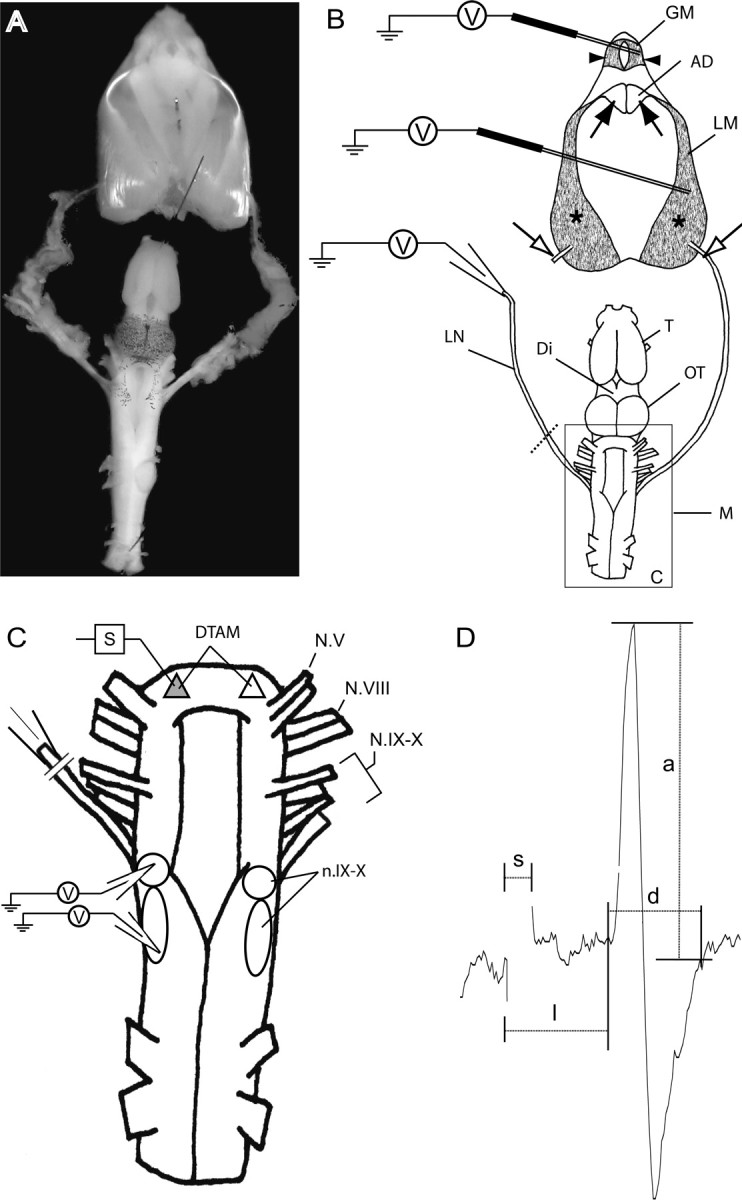
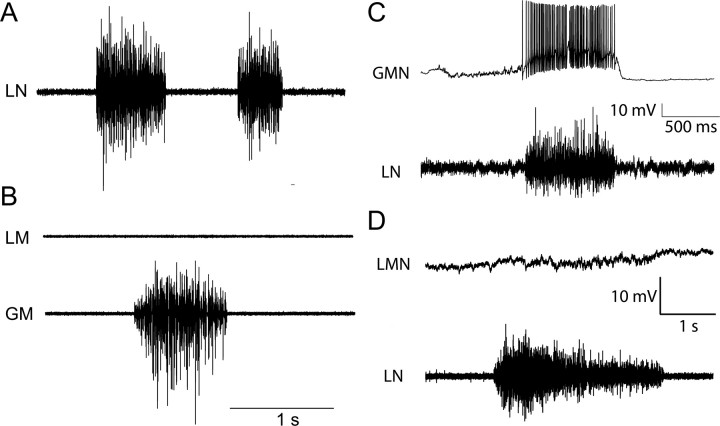
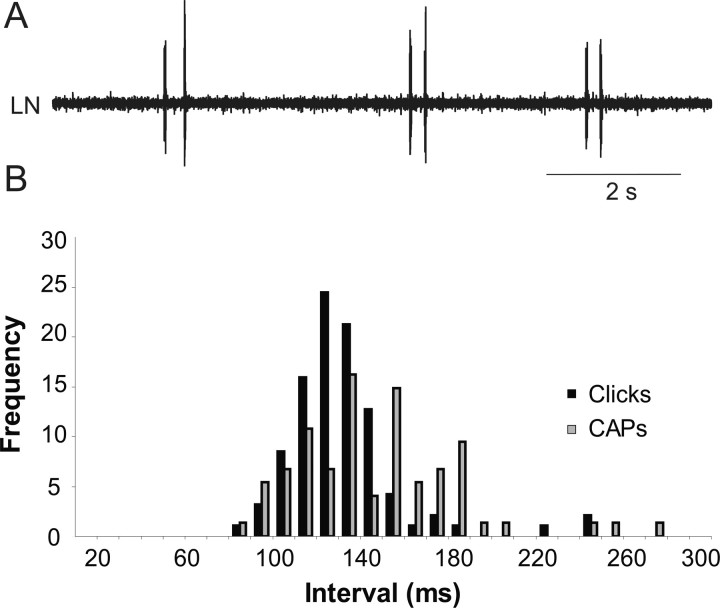
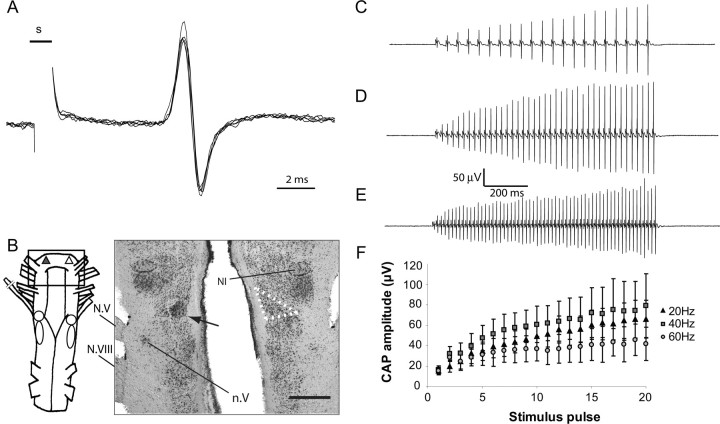
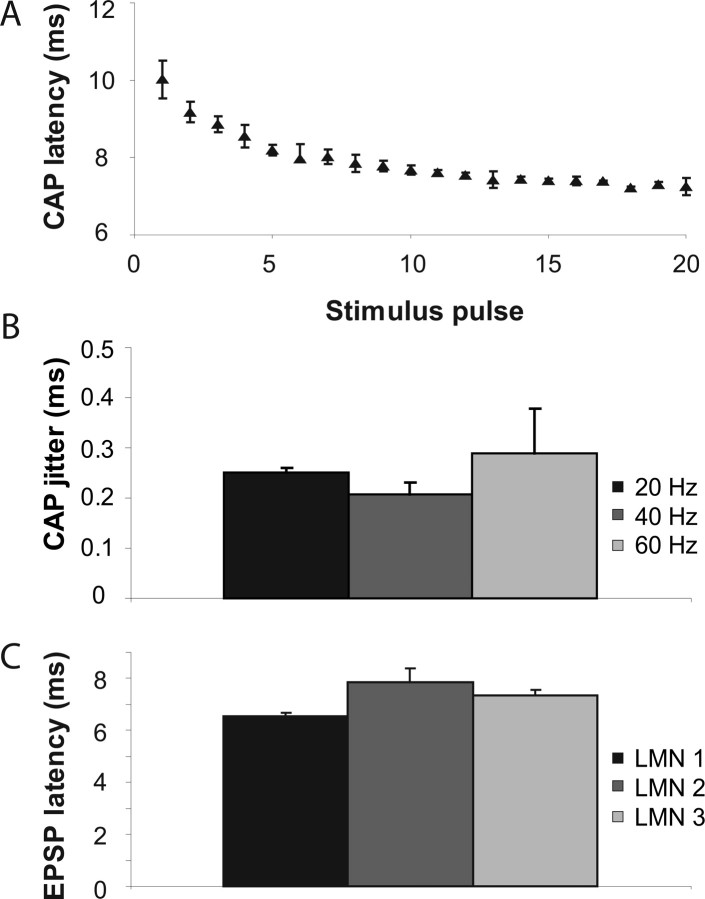
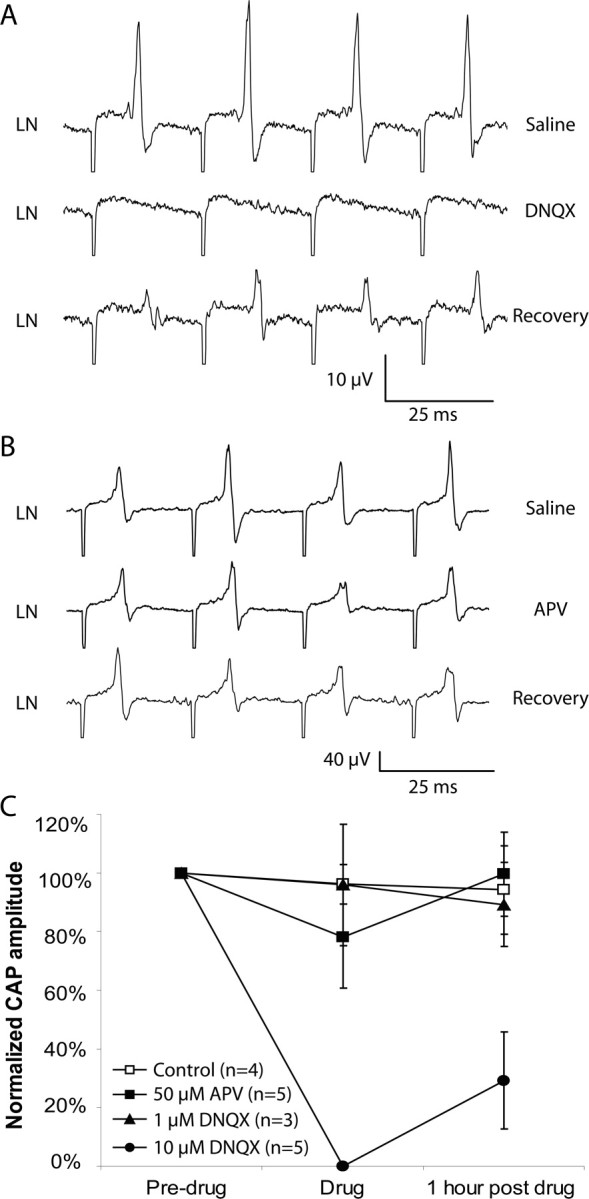
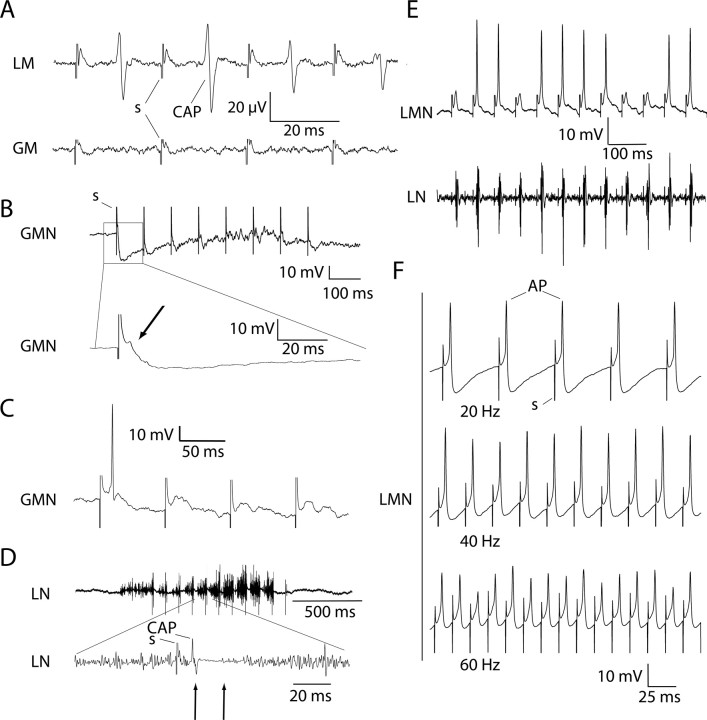
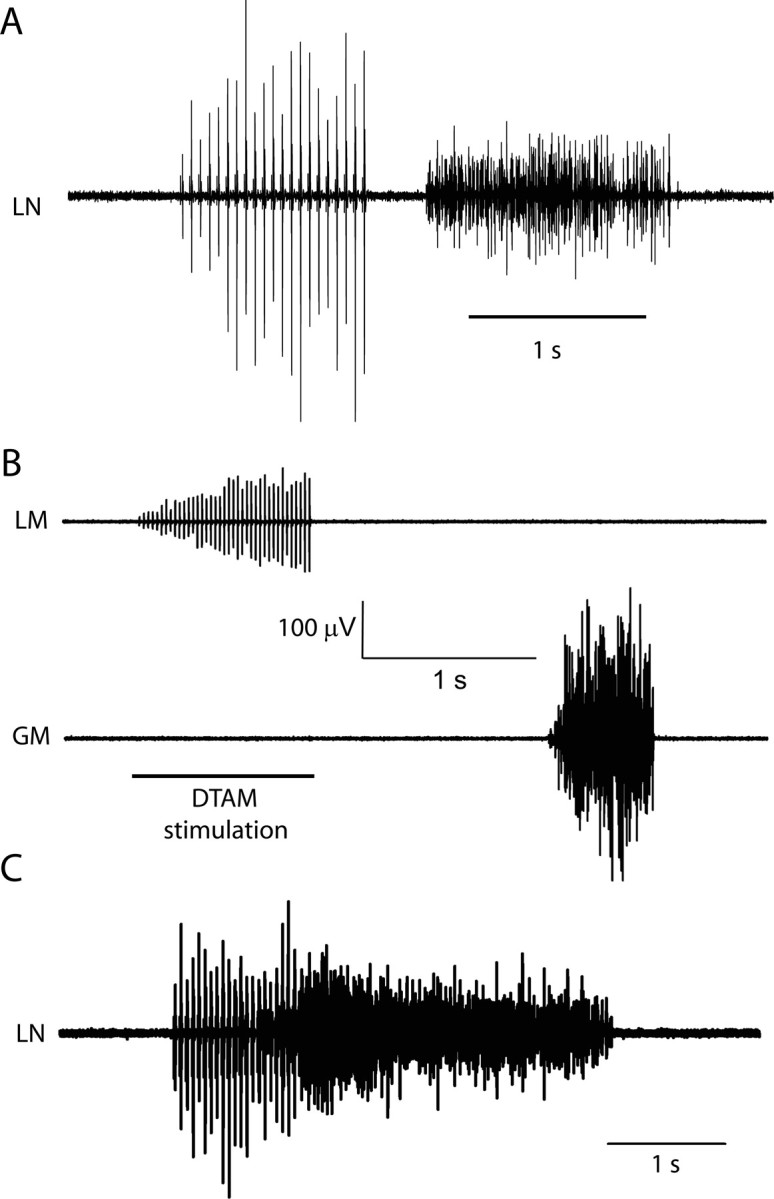
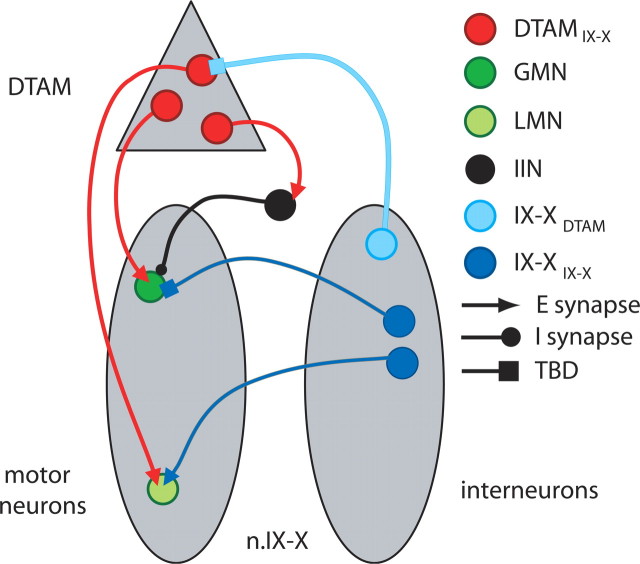
The aquatic frog Xenopus laevis uses a complex vocal repertoire during mating and male-male interactions. Calls are produced without breathing, allowing the frogs to vocalize for long periods underwater. The Xenopus vocal organ, the larynx, is innervated by neurons in cranial motor nucleus (n.) IX-X, which contains both vocal (laryngeal) and respiratory (glottal) motor neurons. The primary descending input to n.IX-X comes from the pretrigeminal nucleus of the dorsal tegmental area of the medulla (DTAM), located in the rostral hindbrain. We wanted to characterize premotor inputs to respiratory and vocal motor neurons and to determine what mechanisms might be involved in regulating two temporally distinct rhythmic behaviors: breathing and calling. Using isolated brain and larynx preparations, we recorded extracellular activity from the laryngeal nerve and muscles and intracellular activity in laryngeal and glottal motor neurons. Spontaneous nerve activities mimicking respiratory and vocal patterns were observed. DTAM projection neurons (DTAM(IX-X) neurons) provide direct input to glottal and laryngeal motor neurons. Electrical stimulation produced short-latency coordinated activity in the laryngeal nerve. DTAM(IX-X) neurons provide excitatory monosynaptic inputs to laryngeal motor neurons and mixed excitatory and inhibitory inputs to glottal motor neurons. DTAM stimulation also produced a delayed burst of glottal motor neuron activity. Together, our data suggest that neurons in DTAM produce vocal motor output by directly activating laryngeal motor neurons and that DTAM may coordinate vocal and respiratory motor activity.
Figures
References
-
- Evans BJ, Kelley DB, Tinsley RC, Melnick DJ, Cannatella DC. A mitochondrial DNA phylogeny of African clawed frogs: phylogeography and implications for polyploid evolution. Mol Phylogenet Evol. 2004;33:197–213. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources