

Intrinsic mechanisms for adaptive gain rescaling in barrel cortex
- PMID: 18199769
- PMCID: PMC6670359
- DOI: 10.1523/JNEUROSCI.4931-07.2008
Intrinsic mechanisms for adaptive gain rescaling in barrel cortex
Abstract
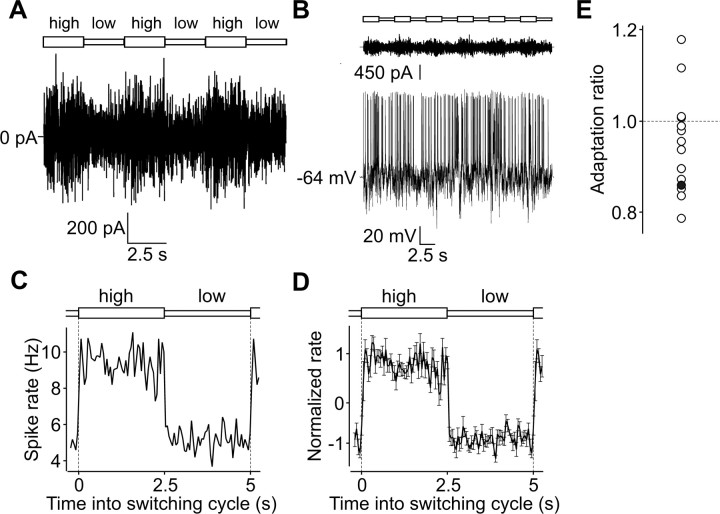
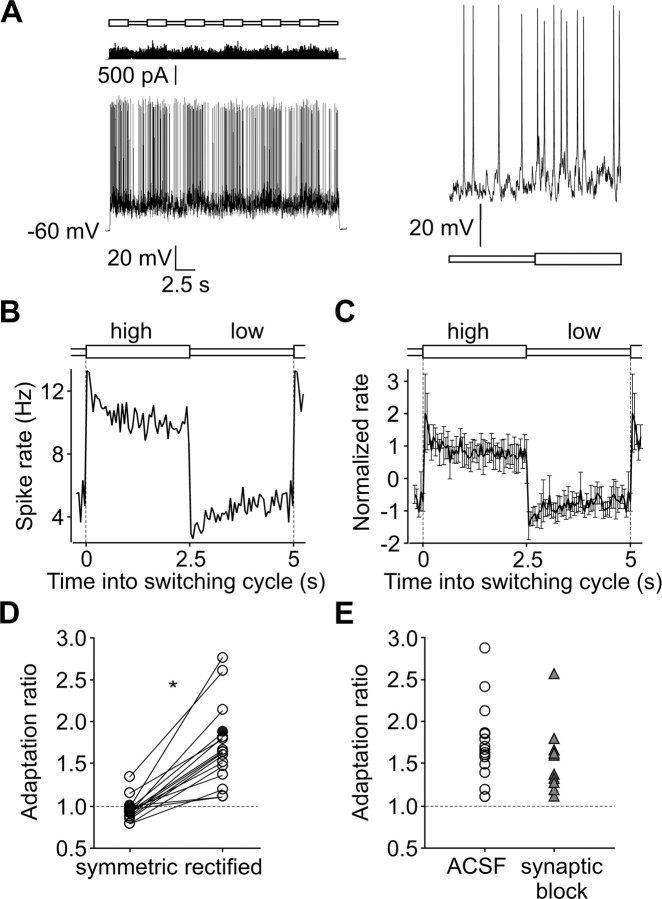
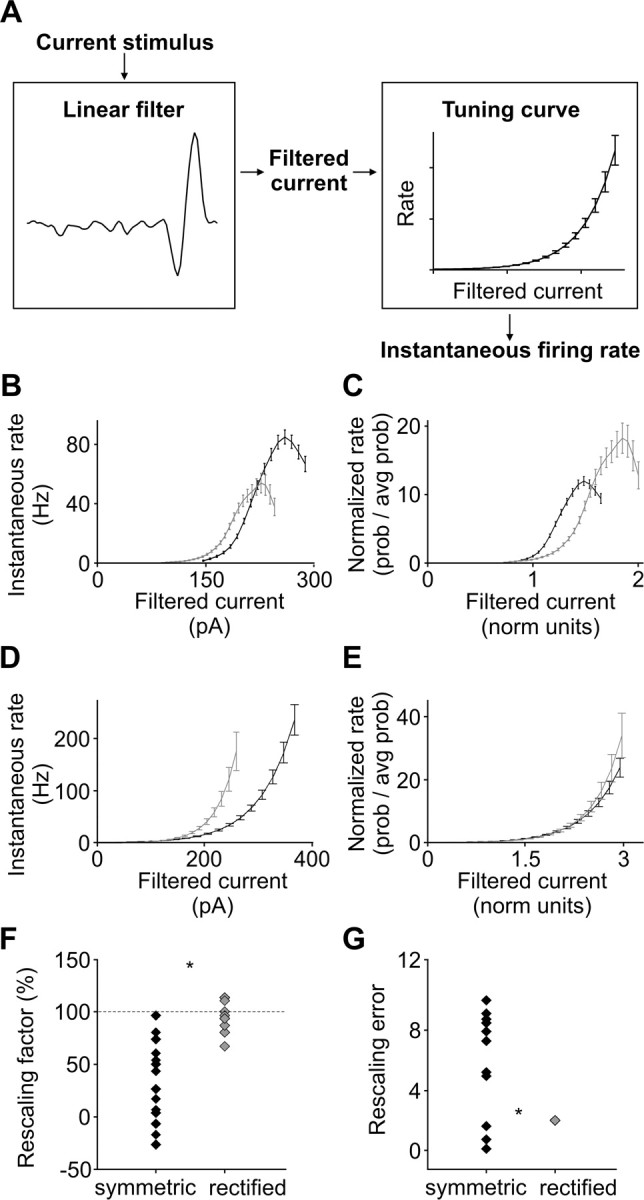
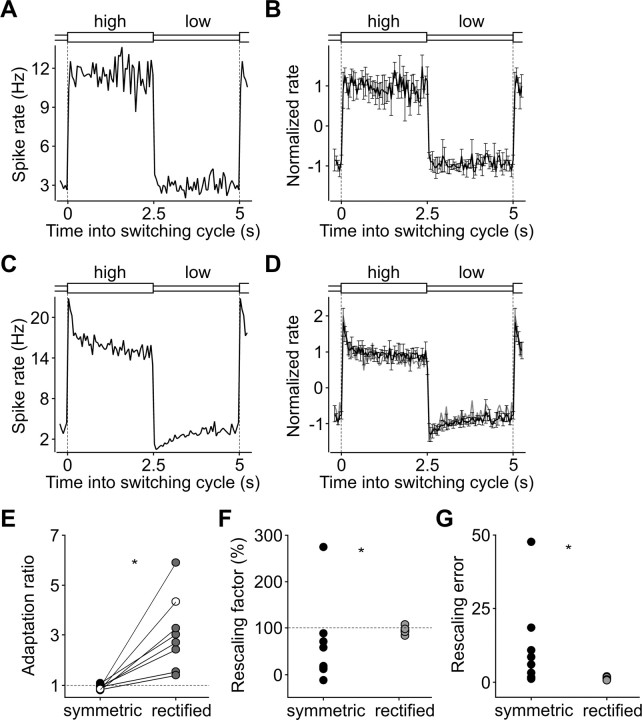
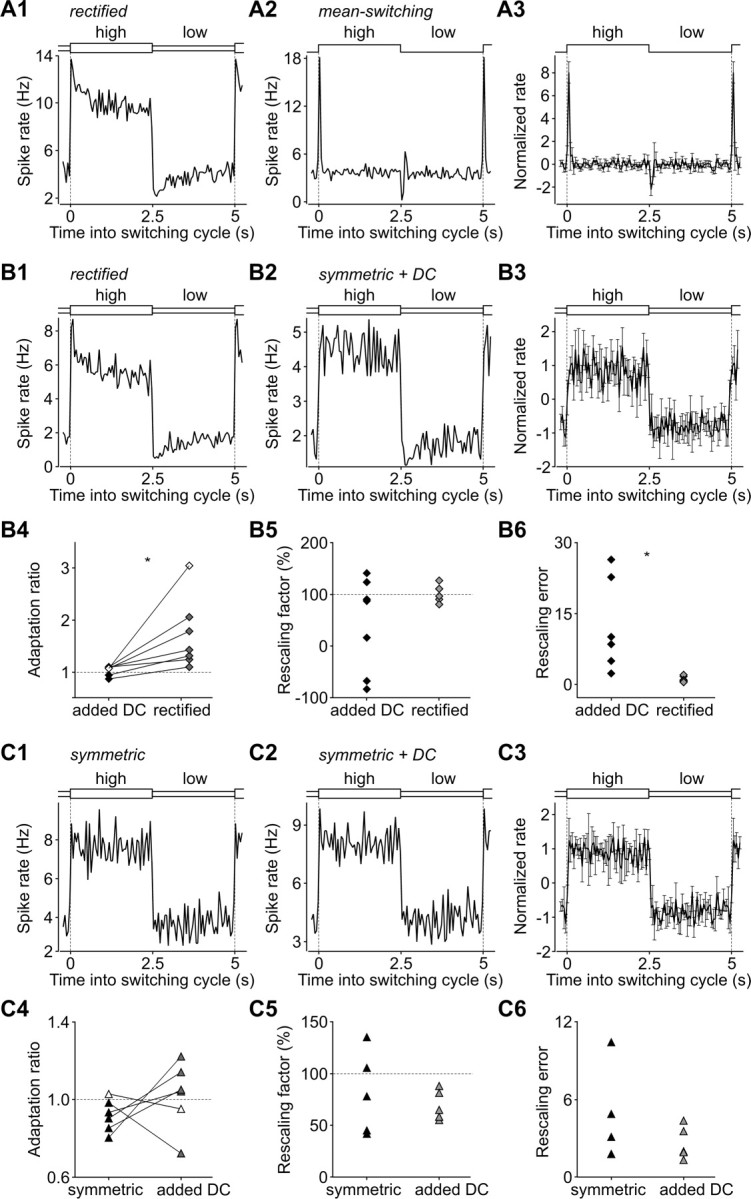
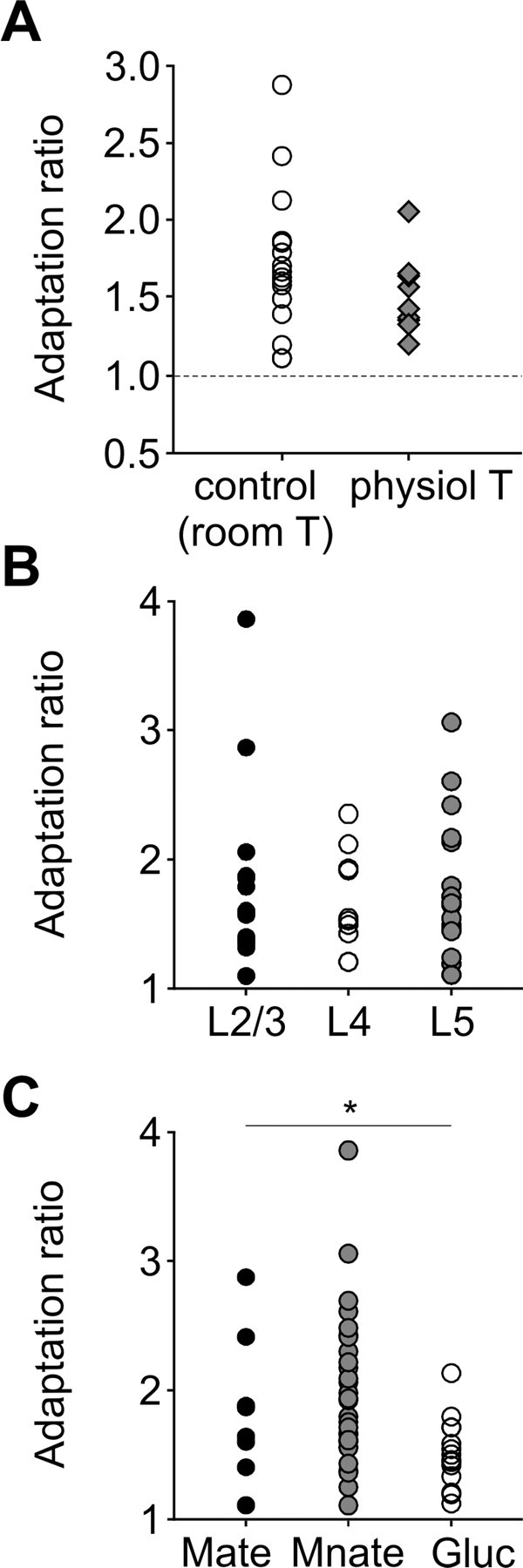
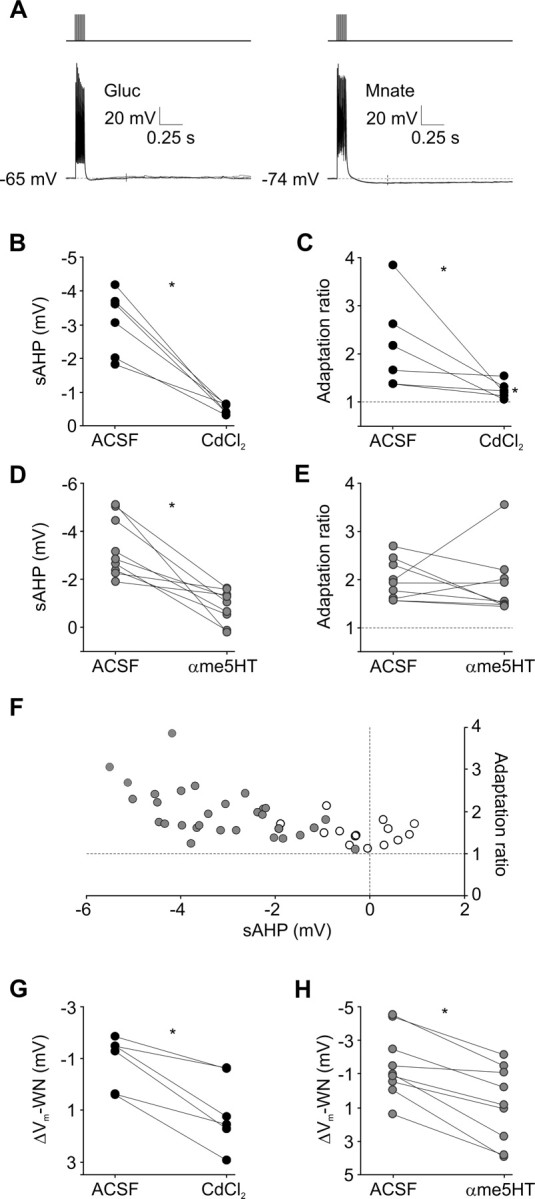
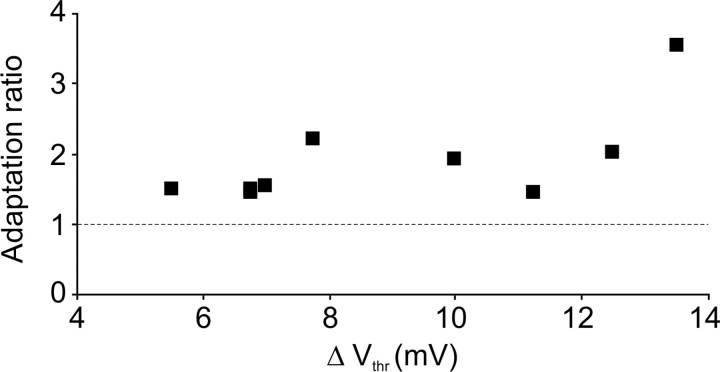
Barrel cortex neuronal responses adapt to changes in the statistics of complex whisker stimuli. This form of adaptation involves an adjustment in the input-output tuning functions of the neurons, such that their gain rescales depending on the range of the current stimulus distribution. Similar phenomena have been observed in other sensory systems, suggesting that adaptive adjustment of responses to ongoing stimulus statistics is an important principle of sensory function. In other systems, adaptation and gain rescaling can depend on intrinsic properties; however, in barrel cortex, whether intrinsic mechanisms can contribute to adaptation to stimulus statistics is unknown. To examine this, we performed whole-cell patch-clamp recordings of pyramidal cells in acute slices while injecting stochastic current stimuli. We induced changes in statistical context by switching across stimulus distributions. The firing rates of neurons adapted in response to changes in stimulus statistics. Adaptation depended on the form of the changes in stimulus distribution: in vivo-like adaptation occurred only for rectified stimuli that maintained neurons in a persistent state of net depolarization. Under these conditions, neurons rescaled the gain of their input-output functions according to the scale of the stimulus distribution, as observed in vivo. This stimulus-specific adaptation was caused by intrinsic properties and correlated strongly with the amplitude of calcium-dependent slow afterhyperpolarizations. Our results suggest that widely expressed intrinsic mechanisms participate in barrel cortex adaptation but that their recruitment is highly stimulus specific.
Figures
Similar articles
-
Shifts in coding properties and maintenance of information transmission during adaptation in barrel cortex.PLoS Biol. 2007 Feb;5(2):e19. doi: 10.1371/journal.pbio.0050019. PLoS Biol. 2007. PMID: 17253902 Free PMC article.
-
Spatial organization of neuronal population responses in layer 2/3 of rat barrel cortex.J Neurosci. 2007 Nov 28;27(48):13316-28. doi: 10.1523/JNEUROSCI.2210-07.2007. J Neurosci. 2007. PMID: 18045926 Free PMC article.
-
Stimulus-specific and stimulus-nonspecific firing synchrony and its modulation by sensory adaptation in the whisker-to-barrel pathway.J Neurophysiol. 2009 May;101(5):2328-38. doi: 10.1152/jn.91151.2008. Epub 2009 Mar 11. J Neurophysiol. 2009. PMID: 19279146 Free PMC article.
-
Neuronal adaptation in the somatosensory system of rodents.Neuroscience. 2017 Feb 20;343:66-76. doi: 10.1016/j.neuroscience.2016.11.043. Epub 2016 Dec 5. Neuroscience. 2017. PMID: 27923742 Review.
-
Sensory Adaptation in the Whisker-Mediated Tactile System: Physiology, Theory, and Function.Front Neurosci. 2021 Oct 29;15:770011. doi: 10.3389/fnins.2021.770011. eCollection 2021. Front Neurosci. 2021. PMID: 34776857 Free PMC article. Review.
Cited by
-
Multiple timescale encoding of slowly varying whisker stimulus envelope in cortical and thalamic neurons in vivo.J Neurosci. 2010 Apr 7;30(14):5071-7. doi: 10.1523/JNEUROSCI.2193-09.2010. J Neurosci. 2010. PMID: 20371827 Free PMC article.
-
Diverse effects of stimulus history in waking mouse auditory cortex.J Neurophysiol. 2017 Aug 1;118(2):1376-1393. doi: 10.1152/jn.00094.2017. Epub 2017 May 31. J Neurophysiol. 2017. PMID: 28566458 Free PMC article.
-
Analysis of Neuronal Spike Trains, Deconstructed.Neuron. 2016 Jul 20;91(2):221-59. doi: 10.1016/j.neuron.2016.05.039. Neuron. 2016. PMID: 27477016 Free PMC article. Review.
-
Postnatal maturation of somatostatin-expressing inhibitory cells in the somatosensory cortex of GIN mice.Front Neural Circuits. 2012 May 31;6:33. doi: 10.3389/fncir.2012.00033. eCollection 2012. Front Neural Circuits. 2012. PMID: 22666189 Free PMC article.
-
Presynaptic Adenosine Receptor-Mediated Regulation of Diverse Thalamocortical Short-Term Plasticity in the Mouse Whisker Pathway.Front Neural Circuits. 2016 Feb 23;10:9. doi: 10.3389/fncir.2016.00009. eCollection 2016. Front Neural Circuits. 2016. PMID: 26941610 Free PMC article.
References
-
- Aguera y Arcas B, Fairhall AL. What causes a neuron to spike? Neural Comput. 2003;15:1789–1807. - PubMed
-
- Aguera y Arcas B, Fairhall AL, Bialek W. Computation in a single neuron: Hodgkin and Huxley revisited. Neural Comput. 2003;15:1715–1749. - PubMed
-
- Ahissar E, Sosnik R, Haidarliu S. Transformation from temporal to rate coding in a somatosensory thalamocortical pathway. Nature. 2000;406:302–306. - PubMed
-
- Ahissar E, Sosnik R, Bagdasarian K, Haidarliu S. Temporal frequency of whisker movement. II. Laminar organization of cortical representations. J Neurophysiol. 2001;86:354–367. - PubMed