

Kalirin-7 is an essential component of both shaft and spine excitatory synapses in hippocampal interneurons
- PMID: 18199770
- PMCID: PMC2570025
- DOI: 10.1523/JNEUROSCI.5283-07.2008
Kalirin-7 is an essential component of both shaft and spine excitatory synapses in hippocampal interneurons
Abstract
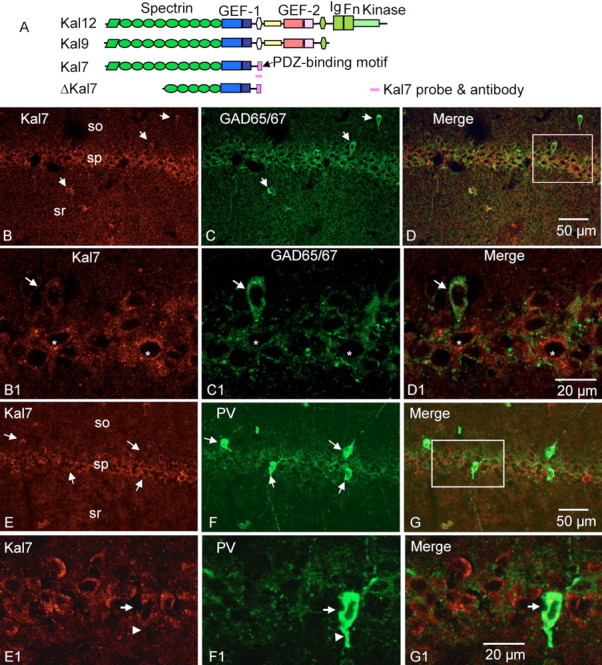
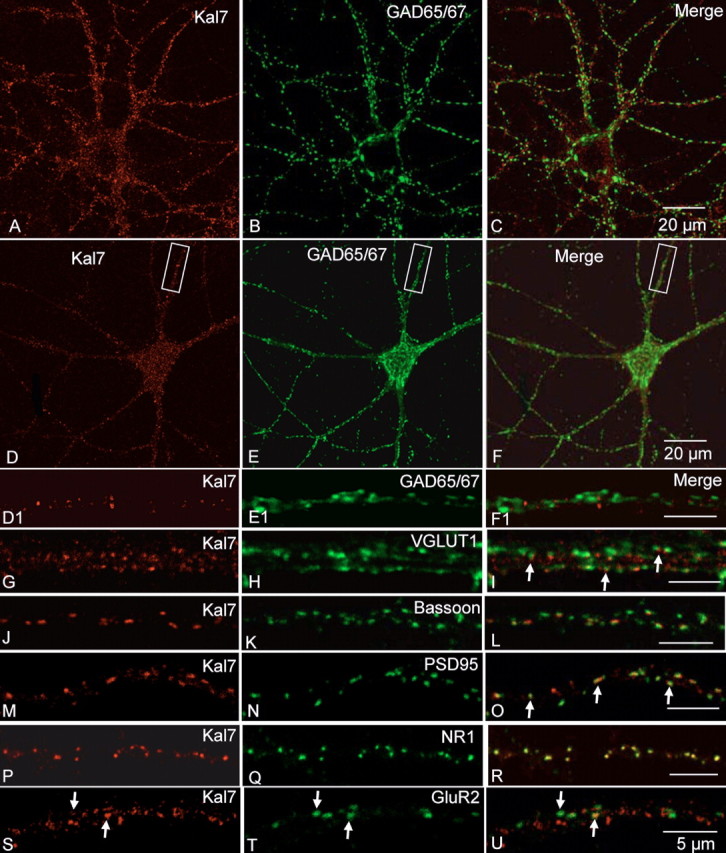
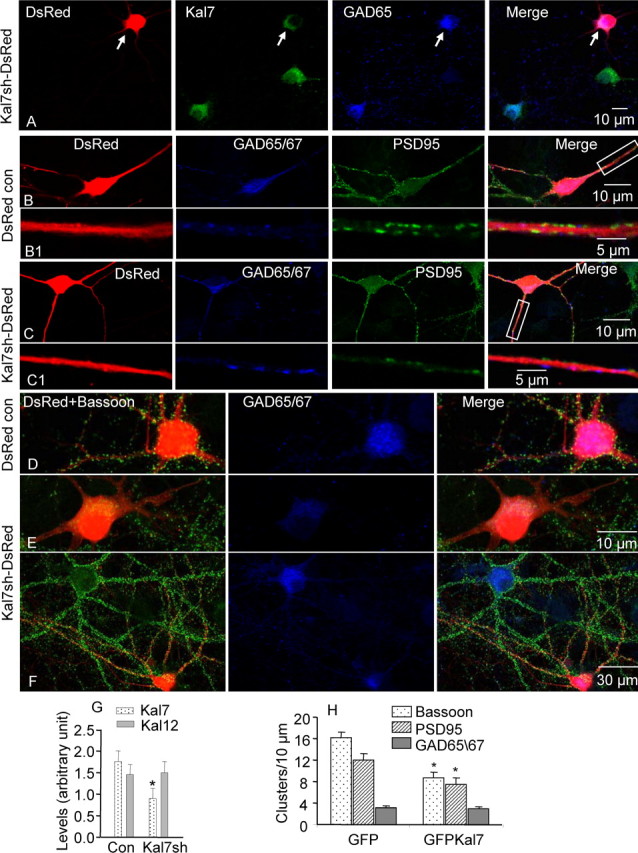
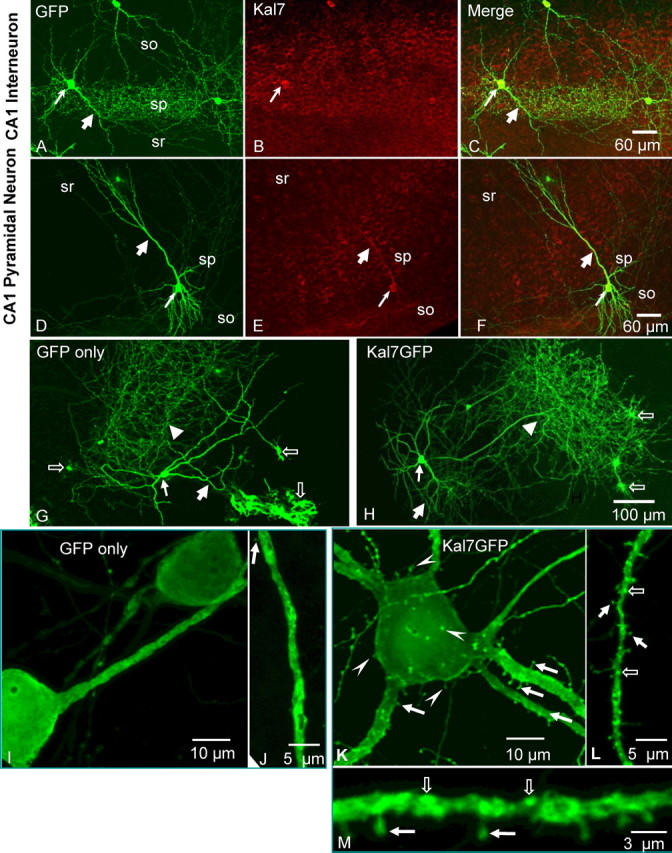
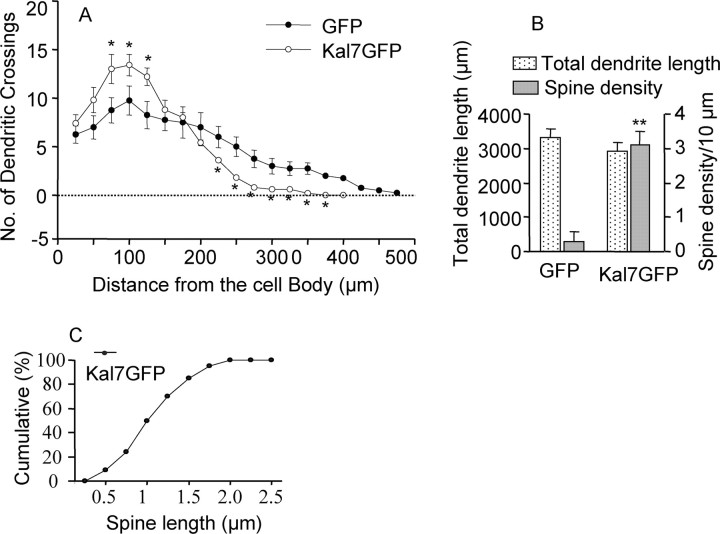
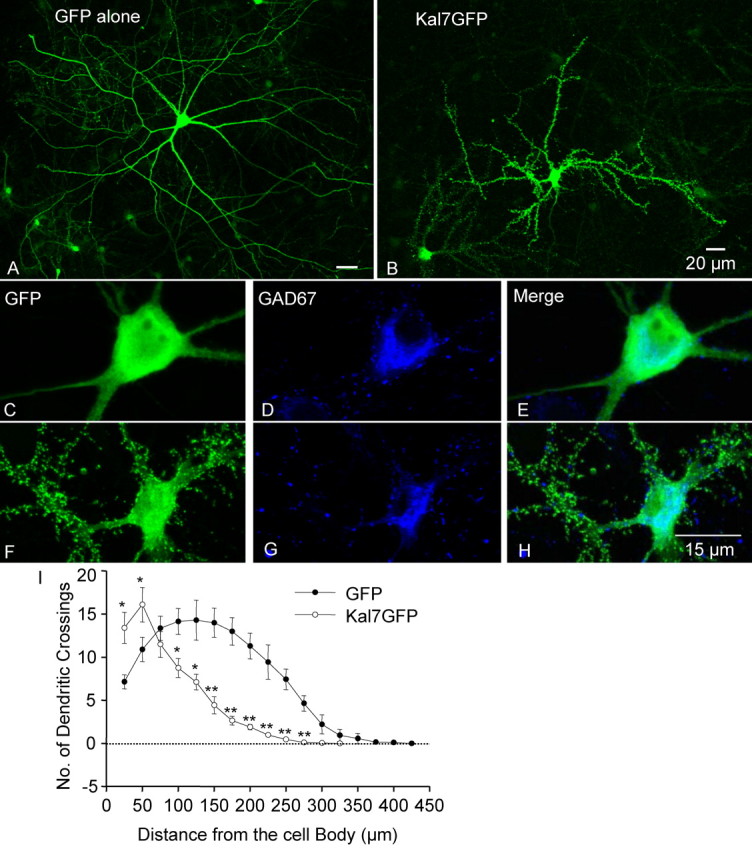
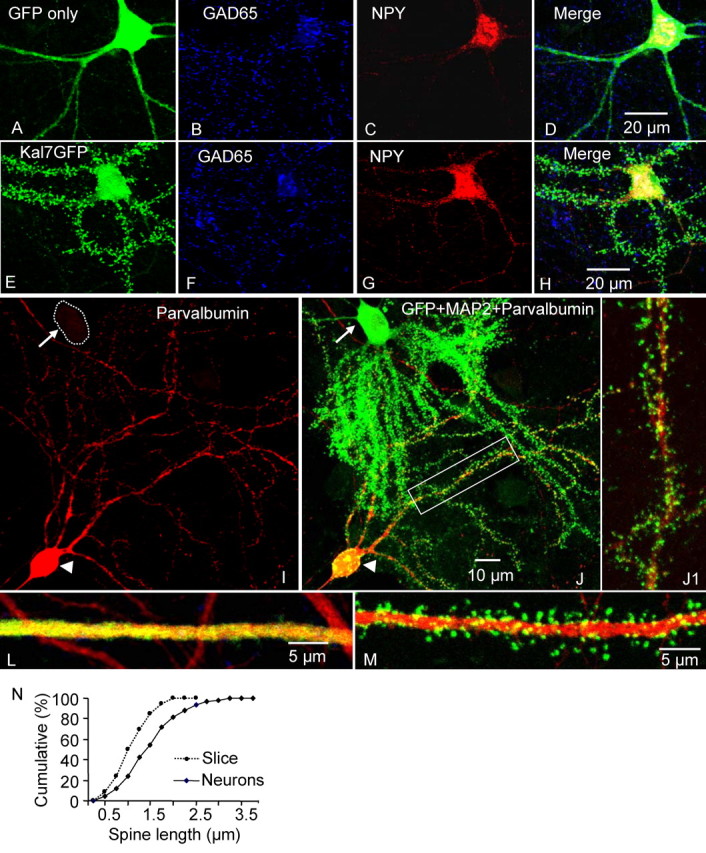
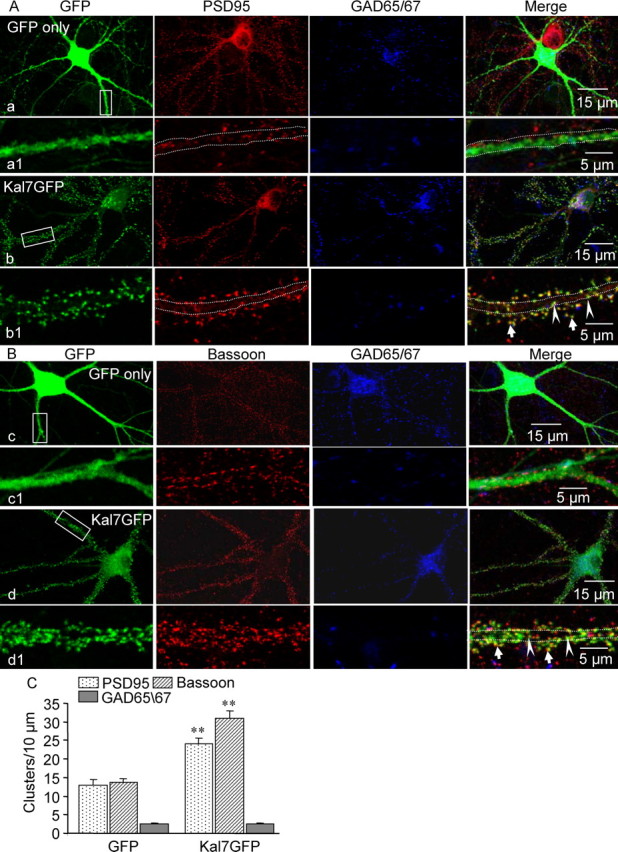
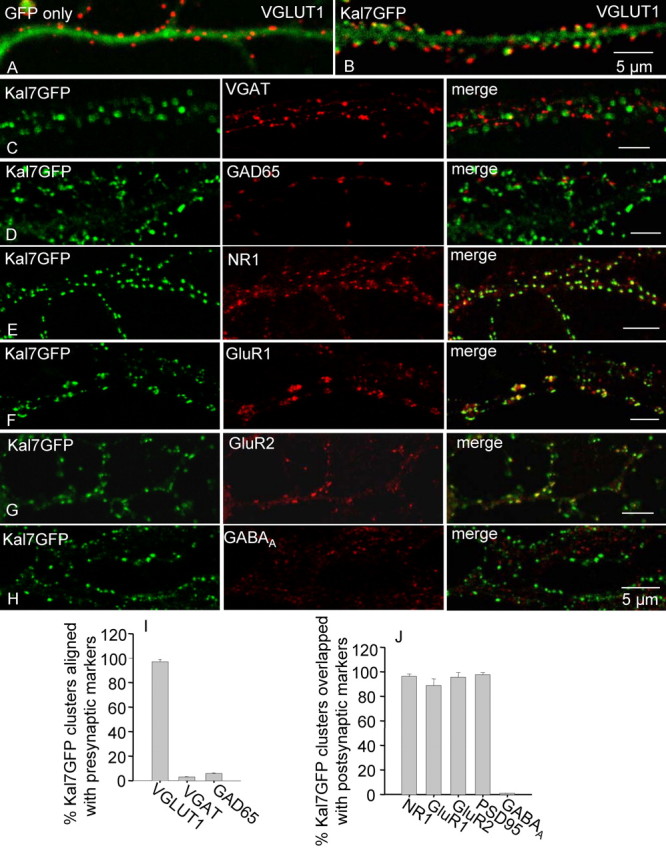
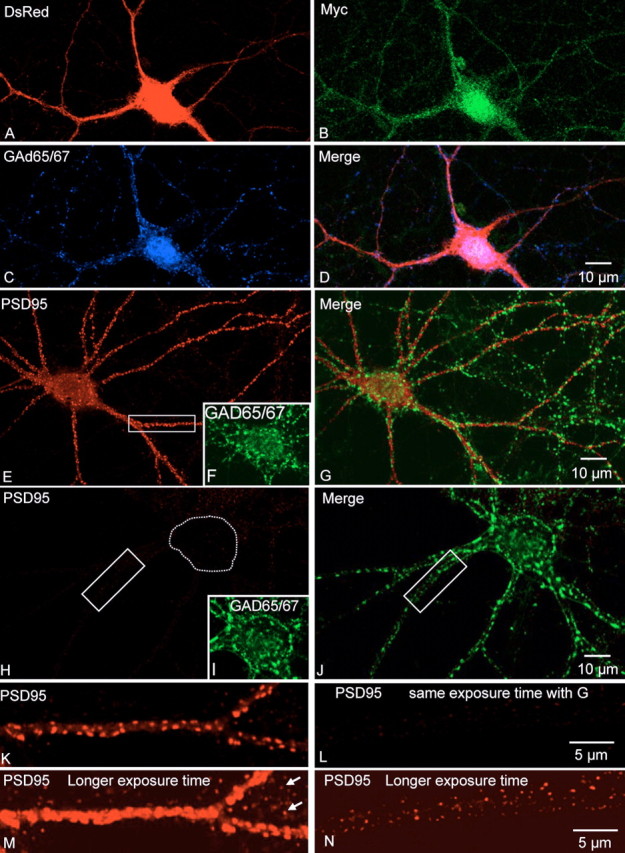
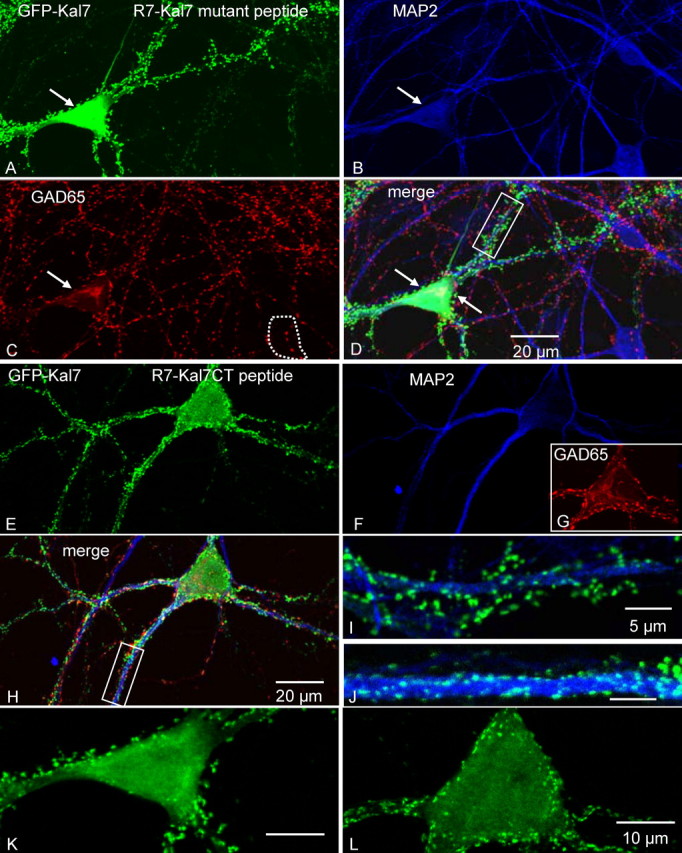
Kalirin, a multifunctional Rho GDP/GTP exchange factor, plays a vital role in cytoskeletal organization, affecting process initiation and outgrowth in neurons. Through alternative splicing, the Kalirin gene generates multiple functionally distinct proteins. Kalirin-7 (Kal7) is the most prevalent isoform in the adult rat hippocampus; it terminates with a postsynaptic density-95 (PSD-95)/Discs large/zona occludens-1 (PDZ) binding motif, is localized to the postsynaptic density, interacts with PSD-95, and causes the formation of dendritic spines when overexpressed in pyramidal neurons. Levels of Kal7 are low in the dendrites of hippocampal aspiny interneurons. In these interneurons, Kal7 is localized to the postsynaptic side of excitatory synapses onto dendritic shafts, overlapping clusters of PSD-95 and NMDA receptor subunit NR1. Selectively decreasing levels of Kal7 decreases the density of PSD-95-positive, bassoon-positive clusters along the dendritic shaft of hippocampal interneurons. Overexpression of Kal7 increases dendritic branching, inducing formation of spine-like structures along the dendrites and on the soma of normally aspiny hippocampal interneurons. Essentially all of the spine-like structures formed in response to Kal7 are apposed to vesicular glutamate transporter 1-positive, bassoon-positive presynaptic endings; GAD-positive, vesicular GABA transporter-positive inhibitory endings are unaffected. Almost every Kal7-positive dendritic cluster contains PSD-95 along with NMDA (NR1) and AMPA (GluR1 and GluR2) receptor subunits. Kal7-induced formation of spine-like structures requires its PDZ binding motif, and interruption of interactions between the PDZ binding motif and its interactors decreases Kal7-induced formation of spine-like structures. Kal7 thus joins Shank3 and GluR2 as molecules with a level of expression at excitatory synapses that titrates the number of dendritic spines.
Figures
Similar articles
-
Kalirin-7, an important component of excitatory synapses, is regulated by estradiol in hippocampal neurons.Hippocampus. 2011 Jun;21(6):661-77. doi: 10.1002/hipo.20780. Epub 2010 Mar 23. Hippocampus. 2011. PMID: 20333733 Free PMC article.
-
Kalirin-7 is required for synaptic structure and function.J Neurosci. 2008 Nov 19;28(47):12368-82. doi: 10.1523/JNEUROSCI.4269-08.2008. J Neurosci. 2008. PMID: 19020030 Free PMC article.
-
Kalirin-7 is a key player in the formation of excitatory synapses in hippocampal neurons.ScientificWorldJournal. 2010 Aug 17;10:1655-66. doi: 10.1100/tsw.2010.148. ScientificWorldJournal. 2010. PMID: 20730383 Free PMC article. Review.
-
Kalirin binds the NR2B subunit of the NMDA receptor, altering its synaptic localization and function.J Neurosci. 2011 Aug 31;31(35):12554-65. doi: 10.1523/JNEUROSCI.3143-11.2011. J Neurosci. 2011. PMID: 21880917 Free PMC article.
-
Kalirin, a key player in synapse formation, is implicated in human diseases.Neural Plast. 2012;2012:728161. doi: 10.1155/2012/728161. Epub 2012 Apr 3. Neural Plast. 2012. PMID: 22548195 Free PMC article. Review.
Cited by
-
Developmental Trajectories of Auditory Cortex Synaptic Structures and Gap-Prepulse Inhibition of Acoustic Startle Between Early Adolescence and Young Adulthood in Mice.Cereb Cortex. 2016 May;26(5):2115-26. doi: 10.1093/cercor/bhv040. Epub 2015 Mar 10. Cereb Cortex. 2016. PMID: 25759333 Free PMC article.
-
Kalirin-9 and Kalirin-12 Play Essential Roles in Dendritic Outgrowth and Branching.Cereb Cortex. 2015 Oct;25(10):3487-501. doi: 10.1093/cercor/bhu182. Epub 2014 Aug 21. Cereb Cortex. 2015. PMID: 25146373 Free PMC article.
-
Synaptic plasticity, a symphony in GEF.ACS Chem Neurosci. 2010 May 19;1(5):348-365. doi: 10.1021/cn100012x. ACS Chem Neurosci. 2010. PMID: 20543890 Free PMC article.
-
Identification of kalirin-7 as a potential post-synaptic density signaling hub.J Proteome Res. 2011 Jun 3;10(6):2828-41. doi: 10.1021/pr200088w. Epub 2011 Apr 25. J Proteome Res. 2011. PMID: 21488700 Free PMC article.
-
Abnormal kalirin signaling in neuropsychiatric disorders.Brain Res Bull. 2014 Apr;103:29-38. doi: 10.1016/j.brainresbull.2013.12.006. Epub 2013 Dec 12. Brain Res Bull. 2014. PMID: 24334022 Free PMC article. Review.
References
-
- Aarts M, Liu Y, Liu L, Besshoh S, Arundine M, Gurd JW, Wang YT, Salter MW, Tymianski M. Treatment of ischemic brain damage by perturbing NMDA receptor- PSD-95 protein interactions. Science. 2002;298:846–850. - PubMed
-
- Anderson TR, Shah PA, Benson DL. Maturation of glutamatergic and GABAergic synapse composition in hippocampal neurons. Neuropharmacology. 2004;47:694–705. - PubMed
-
- Baraban SC, Tallent MK. Interneuron diversity series: interneuronal neuropeptides–endogenous regulators of neuronal excitability. Trends Neurosci. 2004;27:135–142. - PubMed
-
- Benes FM, Berretta S. GABAergic interneurons: implications for understanding schizophrenia and bipolar disorder. Neuropsychopharmacology. 2001;25:1–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources