

HIF-1alpha regulates epithelial inflammation by cell autonomous NFkappaB activation and paracrine stromal remodeling
- PMID: 18199827
- PMCID: PMC2275004
- DOI: 10.1182/blood-2007-10-115758
HIF-1alpha regulates epithelial inflammation by cell autonomous NFkappaB activation and paracrine stromal remodeling
Abstract
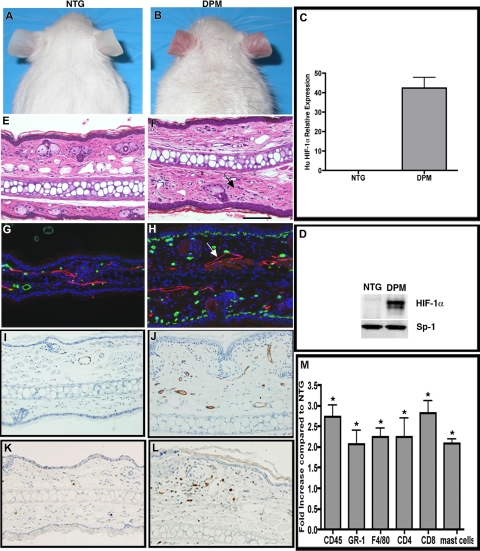
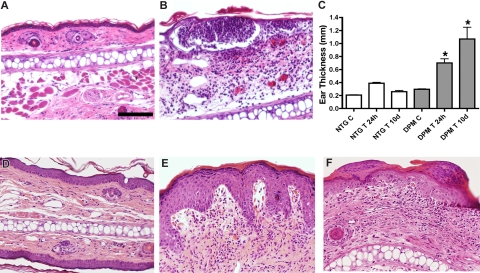
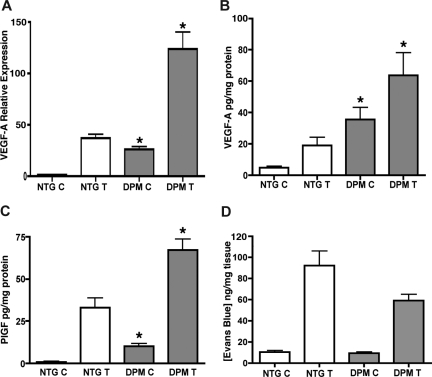
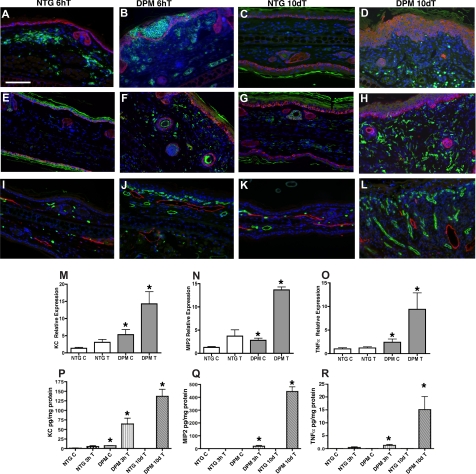
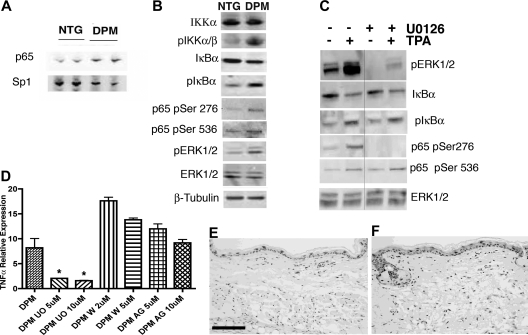
Hypoxia inducible factor-1 (HIF-1) is a master regulatory transcription factor controlling multiple cell-autonomous and non-cell-autonomous processes, such as metabolism, angiogenesis, matrix invasion, and cancer metastasis. Here we used a new line of transgenic mice with constitutive gain of HIF-1 function in basal keratinocytes and demonstrated a signaling pathway from HIF-1 to nuclear factor kappa B (NFkappaB) activation to enhanced epithelial chemokine and cytokine elaboration. This pathway was responsible for a phenotypically silent accumulation of stromal inflammatory cells and a marked inflammatory hypersensitivity to a single 12-O-tetradecanoylphorbol-13-acetate (TPA) challenge. HIF-1-induced NFkappaB activation was composed of 2 elements, IkappaB hyperphosphorylation and phosphorylation of Ser276 on p65, enhancing p65 nuclear localization and transcriptional activity, respectively. NFkappaB transcriptional targets macrophage inflammatory protein-2 (MIP-2/CXCL2/3), keratinocyte chemokine (KC/CXCL1), and tumor necrosis factor [alfa] (TNFalpha) were constitutively up-regulated and further increased after TPA challenge both in cultured keratinocytes and in transgenic mice. Whole animal KC, MIP-2, or TNFalpha immunodepletion each abrogated TPA-induced inflammation, whereas blockade of either VEGF or placenta growth factor (PlGF) signaling did not affect transgenic inflammatory hyper-responsiveness. Thus, epithelial HIF-1 gain of function remodels the local environment by cell-autonomous NFkappaB-mediated chemokine and cytokine secretion, which may be another mechanism by which HIF-1 facilitates either inflammatory diseases or malignant progression.
Figures


References
-
- Karin M, Lawrence T, Nizet V. Innate immunity gone awry: linking microbial infections to chronic inflammation and cancer. Cell. 2006;124:823–835. - PubMed
-
- de Visser KE, Eichten A, Coussens LM. Paradoxical roles of the immune system during cancer development. Nat Rev Cancer. 2006;6:24–37. - PubMed
-
- Zenz R, Eferl R, Kenner L, et al. Psoriasis-like skin disease and arthritis caused by inducible epidermal deletion of Jun proteins. Nature. 2005;437:369–375. - PubMed
-
- Sano S, Chan KS, Kira M, et al. Signal transducer and activator of transcription 3 is a key regulator of keratinocyte survival and proliferation following UV irradiation. Cancer Res. 2005;65:5720–5729. - PubMed
-
- Semenza G. Signal transduction to hypoxia-inducible factor 1. Biochem Pharmacol. 2002;64:993–998. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
