

Contributions of the carboxyl-terminal helical segment to the self-association and lipoprotein preferences of human apolipoprotein E3 and E4 isoforms
- PMID: 18201068
- PMCID: PMC2692928
- DOI: 10.1021/bi701923h
Contributions of the carboxyl-terminal helical segment to the self-association and lipoprotein preferences of human apolipoprotein E3 and E4 isoforms
Abstract
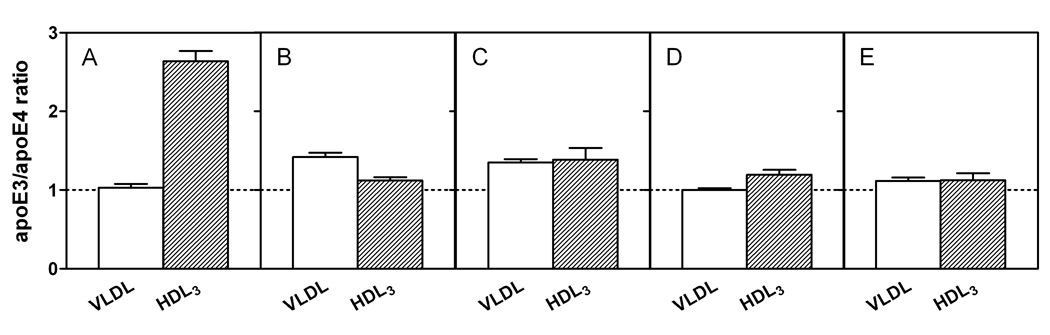
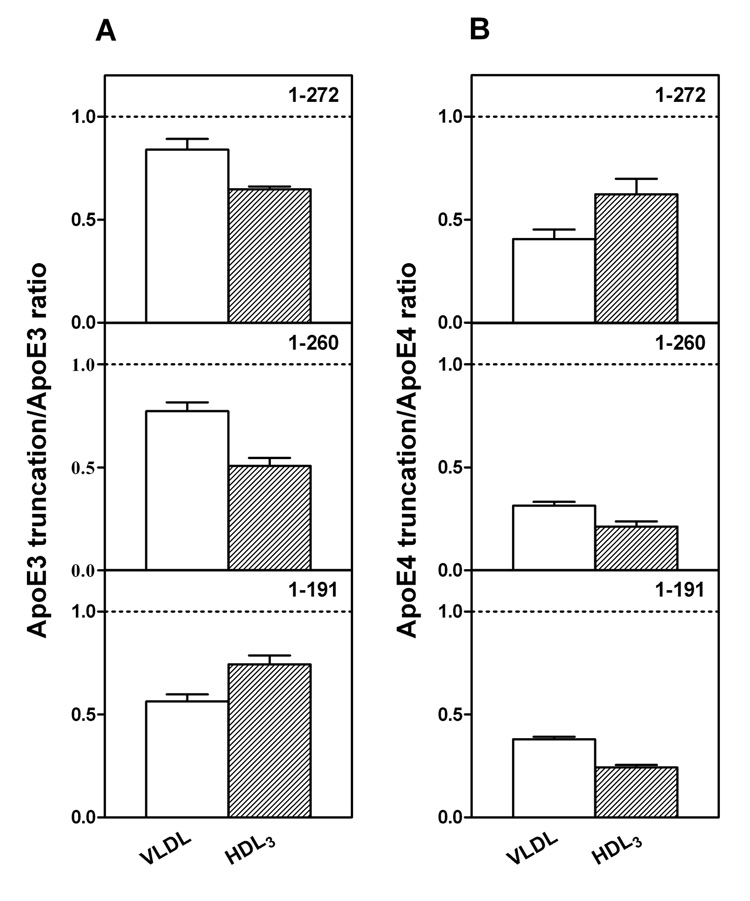
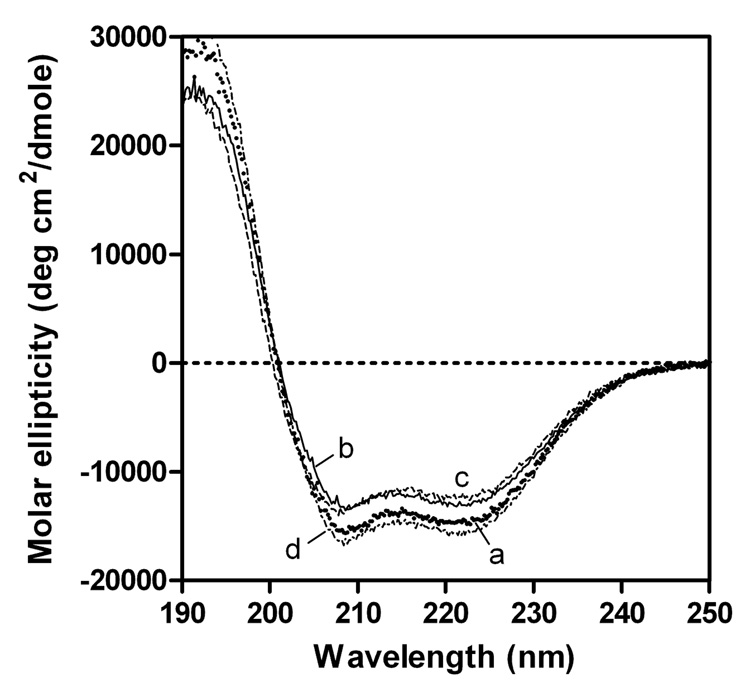
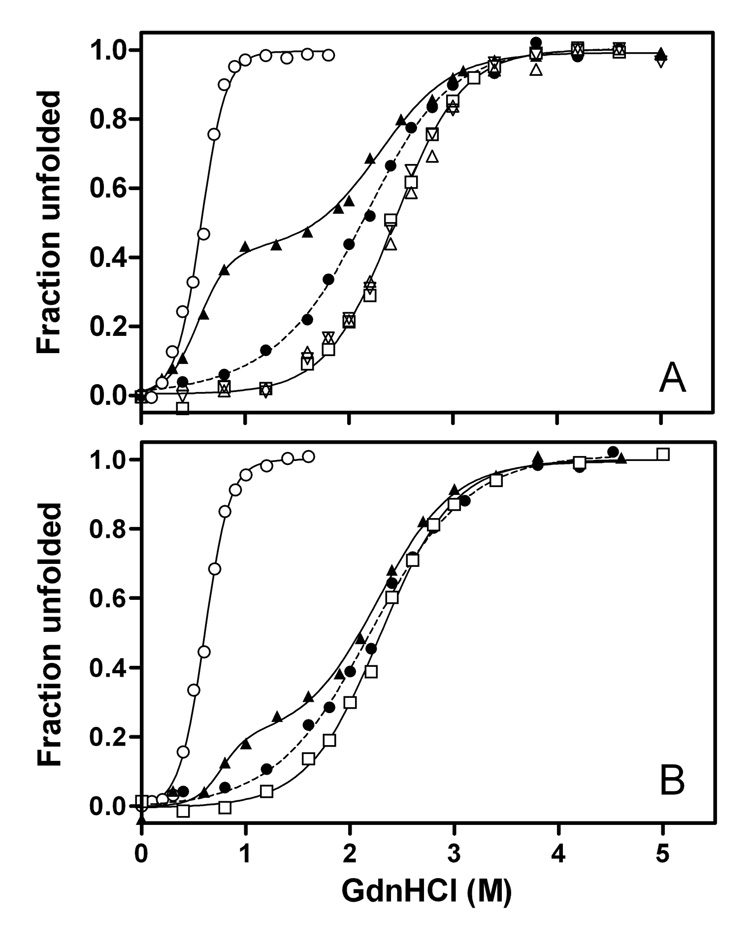
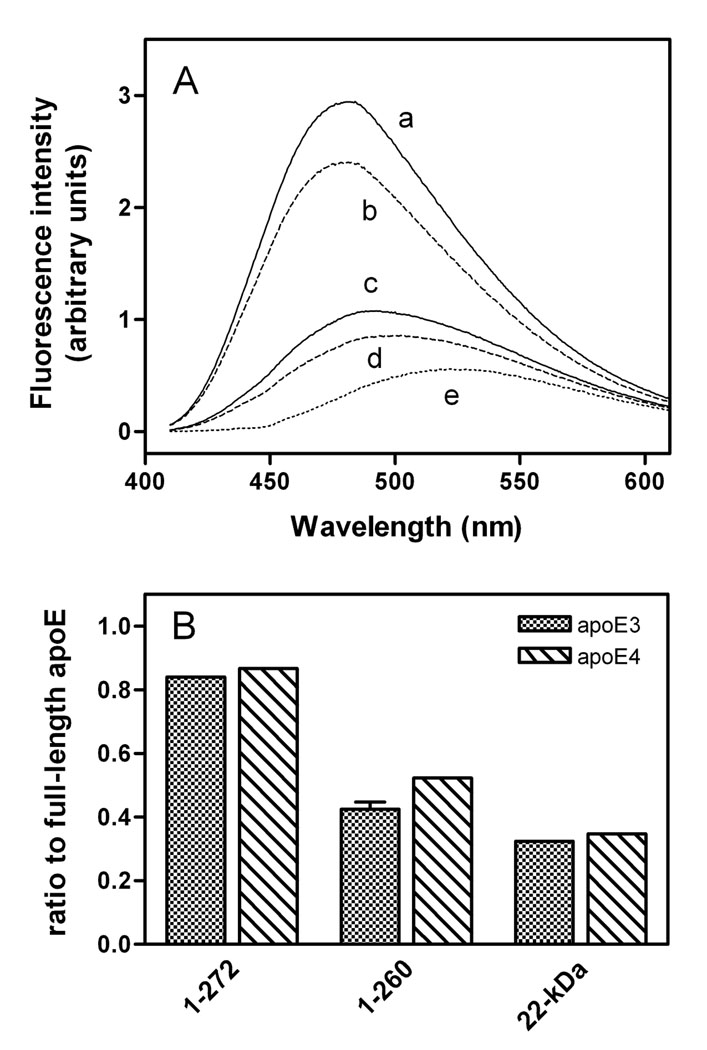
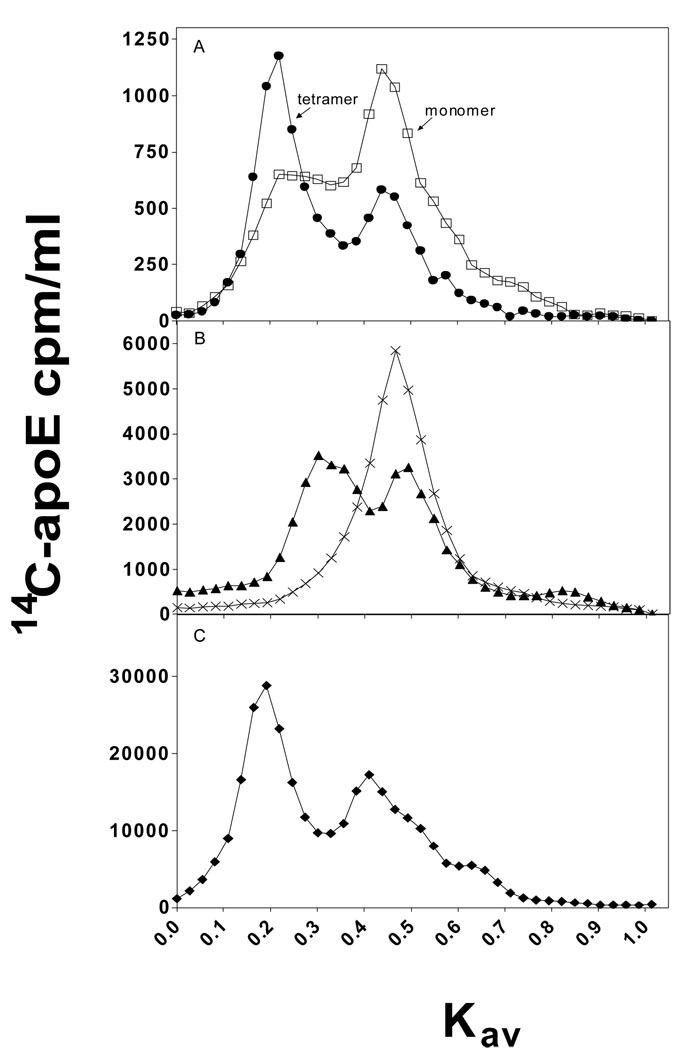
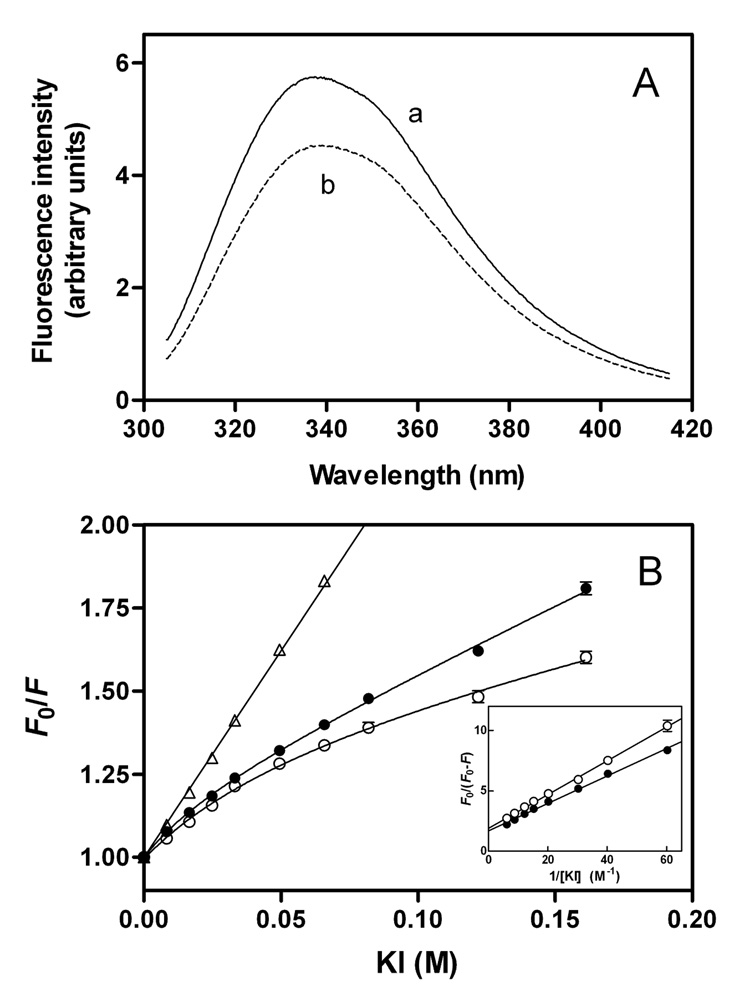
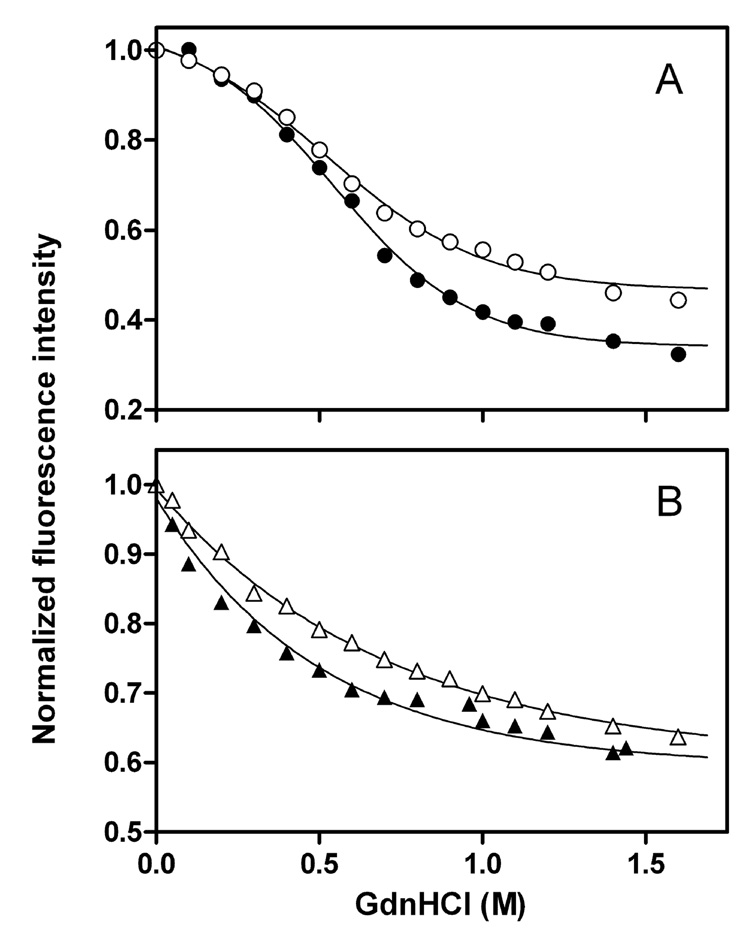
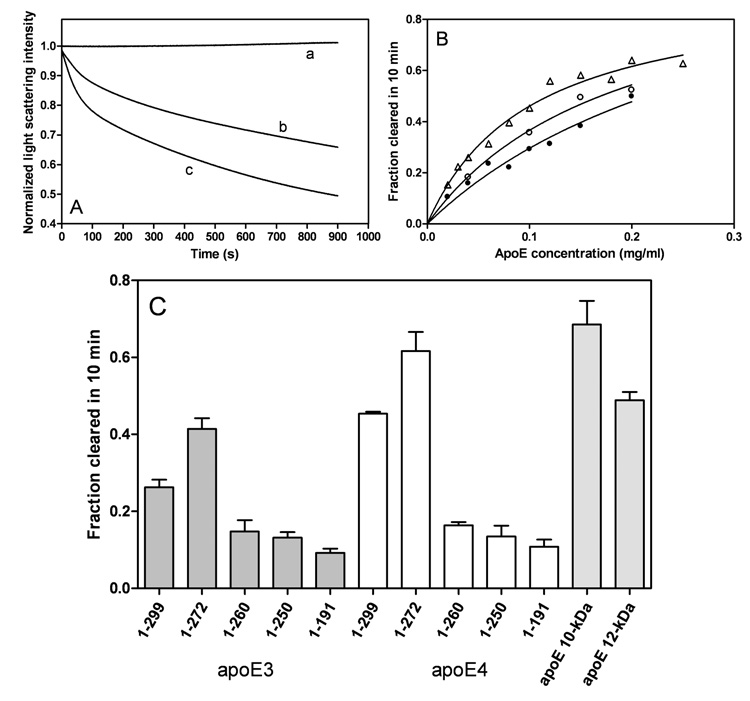
To understand the molecular basis for the different self-association and lipoprotein preferences of apolipoprotein (apo) E isoforms, we compared the effects of progressive truncation of the C-terminal domain in human apoE3 and apoE4 on their lipid-free structure and lipid binding properties. A VLDL/HDL distribution assay demonstrated that apoE3 binds much better than apoE4 to HDL 3, whereas both isoforms bind similarly to VLDL. Removal of the C-terminal helical regions spanning residues 273-299 weakened the ability of both isoforms to bind to lipoproteins; this led to the elimination of the isoform lipoprotein preference, indicating that the C-terminal helices mediate the lipoprotein selectivity of apoE3 and apoE4 isoforms. Gel filtration chromatography experiments demonstrated that the monomer-tetramer distribution is different for the two isoforms with apoE4 being more monomeric than apoE3 and that removal of the C-terminal helices favors the monomeric state in both isoforms. Consistent with this, fluorescence measurements of Trp-264 in single-Trp mutants revealed that the C-terminal domain in apoE4 is less organized and more exposed to the aqueous environment than in apoE3. In addition, the solubilization of dimyristoylphosphatidylcholine multilamellar vesicles is more rapid with apoE4 than with apoE3; removal of the C-terminal helices significantly affected solubilization rates with both isoforms. Taken together, these results indicate that the C-terminal domain is organized differently in apoE3 and apoE4 so that apoE4 self-associates less and binds less than apoE3 to HDL surfaces; these alterations may lead to the pathological sequelae for cardiovascular and neurodegenerative diseases.
Figures
Similar articles
-
Molecular basis for the differences in lipid and lipoprotein binding properties of human apolipoproteins E3 and E4.Biochemistry. 2010 Dec 28;49(51):10881-9. doi: 10.1021/bi1017655. Epub 2010 Dec 3. Biochemistry. 2010. PMID: 21114327 Free PMC article.
-
Fluorescence study of domain structure and lipid interaction of human apolipoproteins E3 and E4.Biochim Biophys Acta. 2014 Dec;1841(12):1716-24. doi: 10.1016/j.bbalip.2014.09.019. Biochim Biophys Acta. 2014. PMID: 25281910 Free PMC article.
-
Conformational analysis of apolipoprotein E3/E4 heteromerization.FEBS J. 2019 May;286(10):1986-1998. doi: 10.1111/febs.14794. Epub 2019 Mar 13. FEBS J. 2019. PMID: 30802357 Free PMC article.
-
Apolipoprotein E: from cardiovascular disease to neurodegenerative disorders.J Mol Med (Berl). 2016 Jul;94(7):739-46. doi: 10.1007/s00109-016-1427-y. Epub 2016 Jun 9. J Mol Med (Berl). 2016. PMID: 27277824 Free PMC article. Review.
-
Apolipoprotein E isoforms and lipoprotein metabolism.IUBMB Life. 2014 Sep;66(9):616-23. doi: 10.1002/iub.1314. IUBMB Life. 2014. PMID: 25328986 Review.
Cited by
-
Fluorescence analysis of the lipid binding-induced conformational change of apolipoprotein E4.Biochemistry. 2012 Jul 17;51(28):5580-8. doi: 10.1021/bi300672s. Epub 2012 Jul 3. Biochemistry. 2012. PMID: 22730894 Free PMC article.
-
Molecular basis for increased risk for late-onset Alzheimer disease due to the naturally occurring L28P mutation in apolipoprotein E4.J Biol Chem. 2014 May 2;289(18):12931-45. doi: 10.1074/jbc.M113.538124. Epub 2014 Mar 18. J Biol Chem. 2014. PMID: 24644280 Free PMC article.
-
Helical structure, stability, and dynamics in human apolipoprotein E3 and E4 by hydrogen exchange and mass spectrometry.Proc Natl Acad Sci U S A. 2017 Jan 31;114(5):968-973. doi: 10.1073/pnas.1617523114. Epub 2017 Jan 17. Proc Natl Acad Sci U S A. 2017. PMID: 28096372 Free PMC article.
-
Role of the N- and C-terminal domains in binding of apolipoprotein E isoforms to heparan sulfate and dermatan sulfate: a surface plasmon resonance study.Biochemistry. 2008 Jun 24;47(25):6702-10. doi: 10.1021/bi8003999. Biochemistry. 2008. PMID: 18507396 Free PMC article.
-
Biochemical and biophysical characterization of recombinant rat apolipoprotein E: similarities to human apolipoprotein E3.Arch Biochem Biophys. 2013 Jan 1;529(1):18-25. doi: 10.1016/j.abb.2012.10.007. Epub 2012 Oct 24. Arch Biochem Biophys. 2013. PMID: 23103361 Free PMC article.
References
-
- Mahley RW. Apolipoprotein E: Cholesterol transport protein with expanding role in cell biology. Science. 1988;240:622–630. - PubMed
-
- Weisgraber KH. Apolipoprotein E: structure-function relationships. Adv. Protein Chem. 1994;45:249–302. - PubMed
-
- Mahley RW, Huang Y. Apolipoprotein E: from atherosclerosis to Alzheimer's disease and beyond. Curr. Opin. Lipidol. 1999;10:207–217. - PubMed
-
- Davignon J. Apolipoprotein E and atherosclerosis: beyond lipid effect. Arterioscler Thromb Vasc Biol. 2005;25:267–269. - PubMed
-
- Mahley RW, Huang Y. Apolipoprotein (apo) E4 and Alzheimer's disease: unique conformational and biophysical properties of apoE4 can modulate neuropathology. Acta Neurol Scand Suppl. 2006;185:8–14. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
