

Effects of long 5' leader sequences on initiation by eukaryotic ribosomes in vitro
- PMID: 1820209
- PMCID: PMC5952206
Effects of long 5' leader sequences on initiation by eukaryotic ribosomes in vitro
Abstract
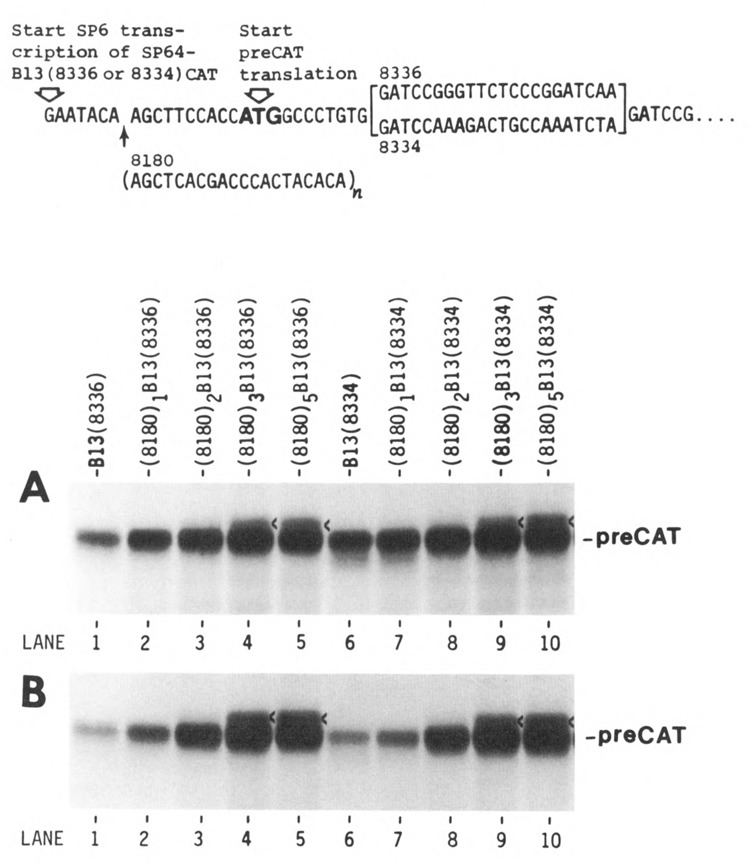
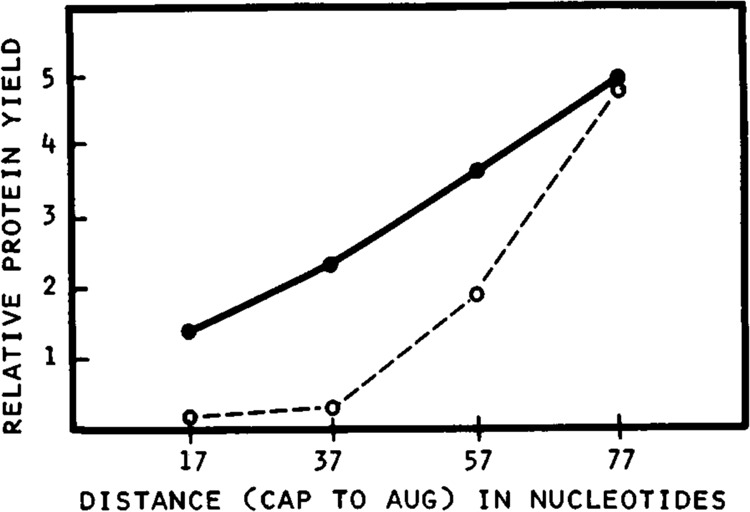
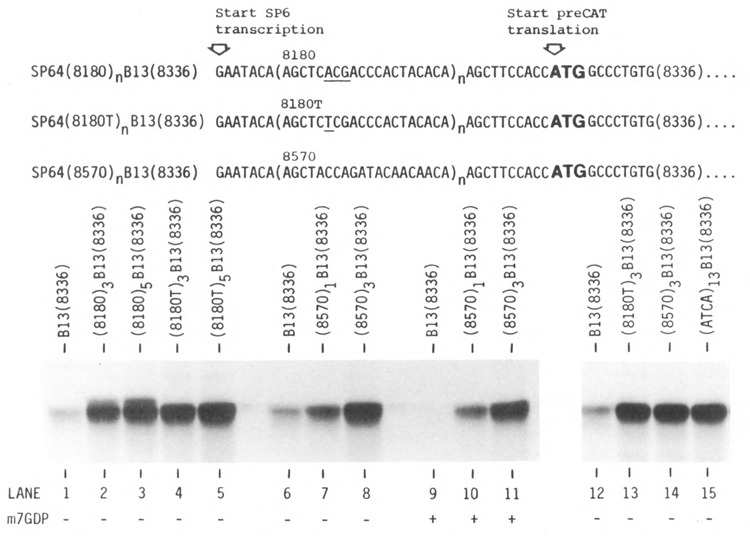
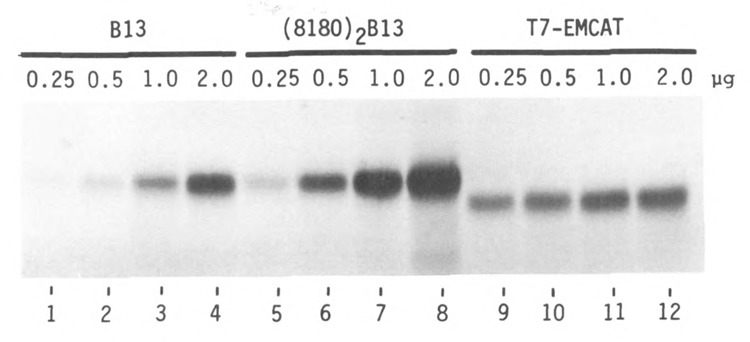
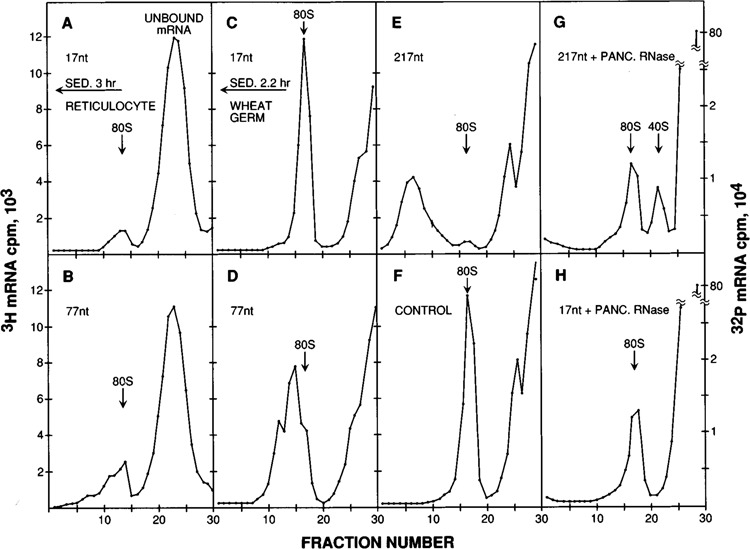
Lengthening the 5' noncoding sequence on SP6-derived transcripts can increase their translational efficiency by an order of magnitude under some conditions of translation in reticulocyte lysates. This effect was observed upon reiterating three different synthetic oligonucleotides, the sequences of which were designed simply to preclude secondary structure. It seems unlikely that such arbitrarily designed sequences are recognized by sequence-specific translational enhancer proteins. Rather, long 5' leader sequences appear to accumulate extra 40S ribosomal subunits, which may account for their translational advantage. The buildup of 40S subunits on long, unstructured leader sequences is predicted by the scanning model for initiation. Leader sequences such as these may be ideal for in vitro expression vectors.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources