

Chromatin-associated genes protect the yeast genome from Ty1 insertional mutagenesis
- PMID: 18202368
- PMCID: PMC2206071
- DOI: 10.1534/genetics.107.082602
Chromatin-associated genes protect the yeast genome from Ty1 insertional mutagenesis
Abstract
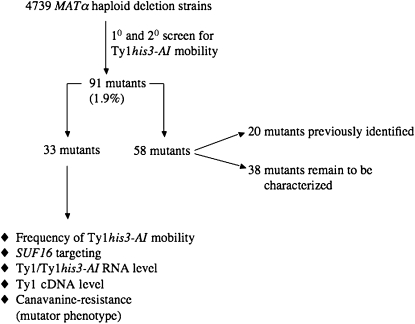
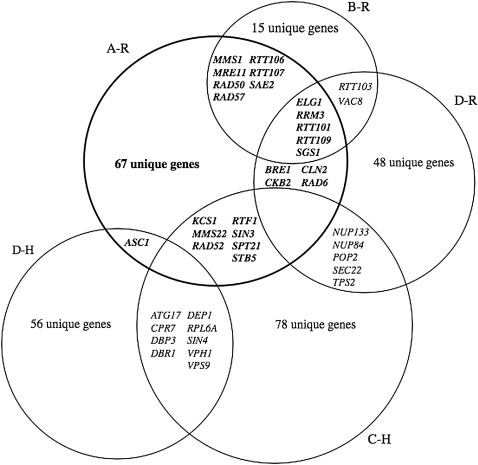
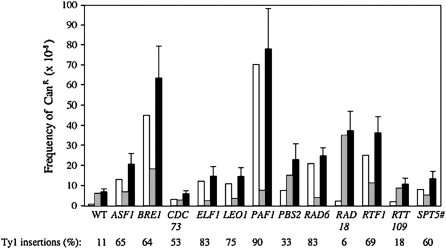
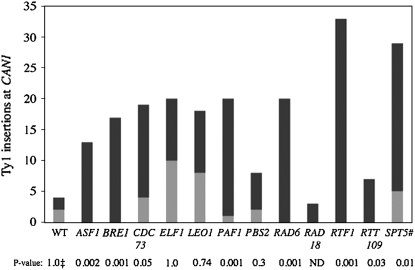
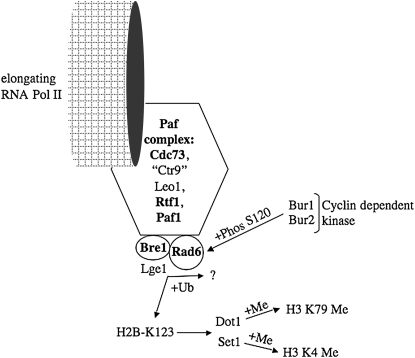
Chromosomal genes modulate Ty retrotransposon movement in the genome of Saccharomyces cerevisiae. We have screened a collection of 4739 deletion mutants to identify those that increase Ty1 mobility (Ty1 restriction genes). Among the 91 identified mutants, 80% encode products involved in nuclear processes such as chromatin structure and function, DNA repair and recombination, and transcription. However, bioinformatic analyses encompassing additional Ty1 and Ty3 screens indicate that 264 unique genes involved in a variety of biological processes affect Ty mobility in yeast. Further characterization of 33 of the mutants identified here show that Ty1 RNA levels increase in 5 mutants and the rest affect mobility post-transcriptionally. RNA and cDNA levels remain unchanged in mutants defective in transcription elongation, including ckb2Delta and elf1Delta, suggesting that Ty1 integration may be more efficient in these strains. Insertion-site preference at the CAN1 locus requires Ty1 restriction genes involved in histone H2B ubiquitination by Paf complex subunit genes, as well as BRE1 and RAD6, histone H3 acetylation by RTT109 and ASF1, and transcription elongation by SPT5. Our results indicate that multiple pathways restrict Ty1 mobility and histone modifications may protect coding regions from insertional mutagenesis.
Figures

Similar articles
-
A nucleosomal surface defines an integration hotspot for the Saccharomyces cerevisiae Ty1 retrotransposon.Genome Res. 2012 Apr;22(4):704-13. doi: 10.1101/gr.129585.111. Epub 2012 Jan 4. Genome Res. 2012. PMID: 22219511 Free PMC article.
-
The yeast Ty1 retrotransposon requires components of the nuclear pore complex for transcription and genomic integration.Nucleic Acids Res. 2018 Apr 20;46(7):3552-3578. doi: 10.1093/nar/gky109. Nucleic Acids Res. 2018. PMID: 29514267 Free PMC article.
-
CRISPR/Transposon gene integration (CRITGI) can manage gene expression in a retrotransposon-dependent manner.Sci Rep. 2019 Oct 25;9(1):15300. doi: 10.1038/s41598-019-51891-6. Sci Rep. 2019. PMID: 31653950 Free PMC article.
-
Happy together: the life and times of Ty retrotransposons and their hosts.Cytogenet Genome Res. 2005;110(1-4):70-90. doi: 10.1159/000084940. Cytogenet Genome Res. 2005. PMID: 16093660 Review.
-
Ty1 integrase overexpression leads to integration of non-Ty1 DNA fragments into the genome of Saccharomyces cerevisiae.Mol Genet Genomics. 2010 Oct;284(4):231-42. doi: 10.1007/s00438-010-0561-4. Epub 2010 Jul 31. Mol Genet Genomics. 2010. PMID: 20677012 Free PMC article. Review.
Cited by
-
A role for the budding yeast separase, Esp1, in Ty1 element retrotransposition.PLoS Genet. 2015 Mar 30;11(3):e1005109. doi: 10.1371/journal.pgen.1005109. eCollection 2015 Mar. PLoS Genet. 2015. PMID: 25822502 Free PMC article.
-
Mapping genomic hotspots of DNA damage by a single-strand-DNA-compatible and strand-specific ChIP-seq method.Genome Res. 2013 Apr;23(4):705-15. doi: 10.1101/gr.146357.112. Epub 2012 Dec 17. Genome Res. 2013. PMID: 23249883 Free PMC article.
-
Control of yeast retrotransposons mediated through nucleoporin evolution.PLoS Genet. 2018 Apr 25;14(4):e1007325. doi: 10.1371/journal.pgen.1007325. eCollection 2018 Apr. PLoS Genet. 2018. PMID: 29694349 Free PMC article.
-
mChIP-KAT-MS, a method to map protein interactions and acetylation sites for lysine acetyltransferases.Proc Natl Acad Sci U S A. 2013 Apr 23;110(17):E1641-50. doi: 10.1073/pnas.1218515110. Epub 2013 Apr 9. Proc Natl Acad Sci U S A. 2013. PMID: 23572591 Free PMC article.
-
Genome-wide distribution of RNA-DNA hybrids identifies RNase H targets in tRNA genes, retrotransposons and mitochondria.PLoS Genet. 2014 Oct 30;10(10):e1004716. doi: 10.1371/journal.pgen.1004716. eCollection 2014 Oct. PLoS Genet. 2014. PMID: 25357144 Free PMC article.
References
-
- Ahmed, K., D. A. Gerber and C. Cochet, 2002. Joining the cell survival squad: an emerging role for protein kinase CK2. Trends Cell Biol. 12: 226–230. - PubMed
-
- Alcazar-Roman, A. R., E. J. Tran, S. Guo and S. R. Wente, 2006. Inositol hexakisphosphate and Gle1 activate the DEAD-box protein Dbp5 for nuclear mRNA export. Nat. Cell Biol. 8: 711–716. - PubMed
-
- Auesukaree, C., H. Tochio, M. Shirakawa, Y. Kaneko and S. Harashima, 2005. Plc1p, Arg82p, and Kcs1p, enzymes involved in inositol pyrophosphate synthesis, are essential for phosphate regulation and polyphosphate accumulation in Saccharomyces cerevisiae. J. Biol. Chem. 280: 25127–25133. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
