

A gain-of-function suppressor screen for genes involved in dorsal-ventral boundary formation in the Drosophila wing
- PMID: 18202376
- PMCID: PMC2206080
- DOI: 10.1534/genetics.107.081869
A gain-of-function suppressor screen for genes involved in dorsal-ventral boundary formation in the Drosophila wing
Abstract
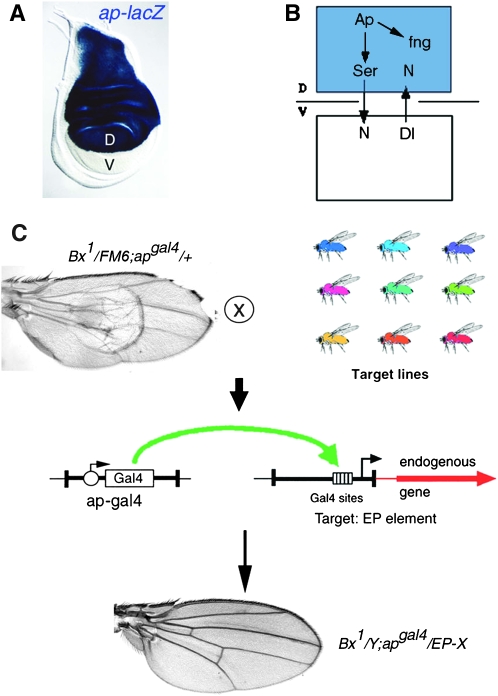
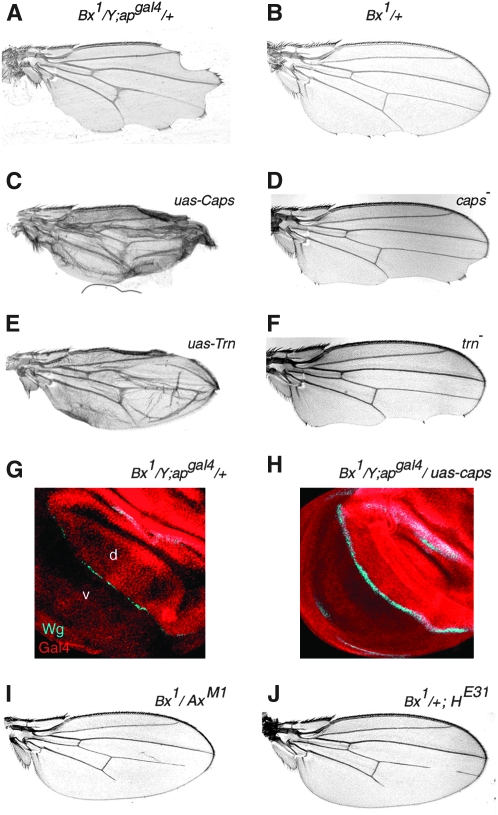
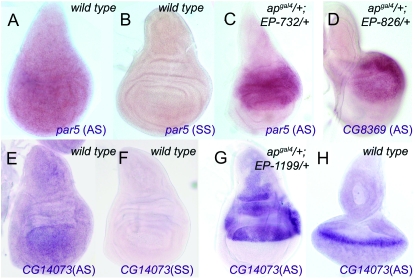
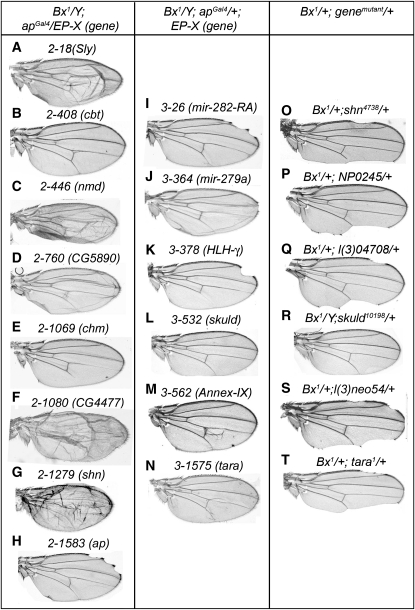
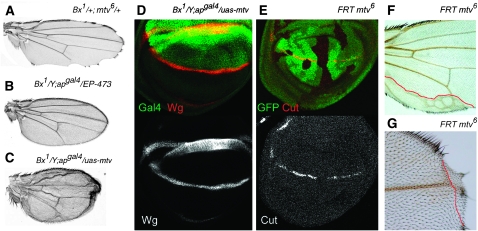
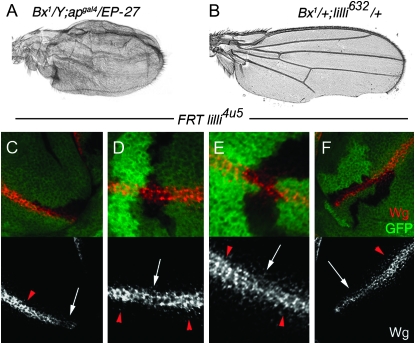
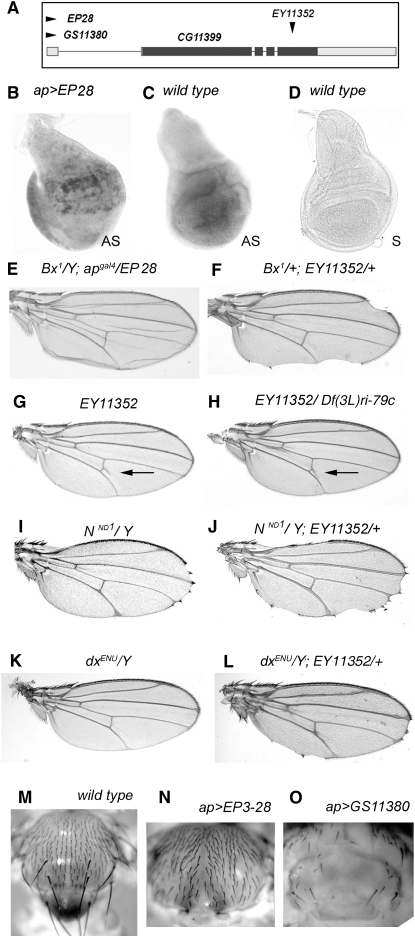
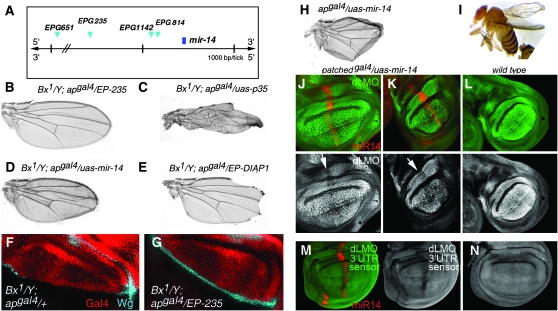
The Drosophila wing primordium is subdivided into a dorsal (D) and a ventral (V) compartment by the activity of the LIM-homeodomain protein Apterous in D cells. Cell interactions between D and V cells induce the activation of Notch at the DV boundary. Notch is required for the maintenance of the compartment boundary and the growth of the wing primordium. Beadex, a gain-of-function allele of dLMO, results in increased levels of dLMO protein, which interferes with the activity of Apterous and results in defects in DV axis formation. We performed a gain-of-function enhancer-promoter (EP) screen to search for suppressors of Beadex when overexpressed in D cells. We identified 53 lines corresponding to 35 genes. Loci encoding for micro-RNAs and proteins involved in chromatin organization, transcriptional control, and vesicle trafficking were characterized in the context of dLMO activity and DV boundary formation. Our results indicate that a gain-of-function genetic screen in a sensitized background, as opposed to classical loss-of-function-based screenings, is a very efficient way to identify redundant genes involved in a developmental process.
Figures
References
-
- Adachi-Yamada, T., K. Fujimura-Kamada, Y. Nishida and K. Matsumoto, 1999. Distortion of proximodistal information causes JNK-dependent apoptosis in Drosophila wing. Nature 400: 166–169. - PubMed
-
- Aurora, K., H. Dai, S. G. Kazuko, J. Jamal, M. B. O'Connor et al., 1995. The Drosophila schnurri gene acts in the Dpp/TGFβ signalling pathway and encodes a transcription factor homologous to the human MBP family. Cell 81: 781–790. - PubMed
-
- Bang, A. G., A. M. Bailey and J. W. Posakony, 1995. Hairless promotes stable commitment to the sensory organ precursor cell fate by negatively regulating the activity of the Notch signaling pathway. Dev. Biol. 172: 479–494. - PubMed
-
- Barabasi, A. L., and Z. N. Oltvai, 2004. Network biology: understanding the cell's functional organization. Nat. Rev. Genet. 5: 101–113. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
