

Cyclic dermal BMP signalling regulates stem cell activation during hair regeneration
- PMID: 18202659
- PMCID: PMC2696201
- DOI: 10.1038/nature06457
Cyclic dermal BMP signalling regulates stem cell activation during hair regeneration
Abstract
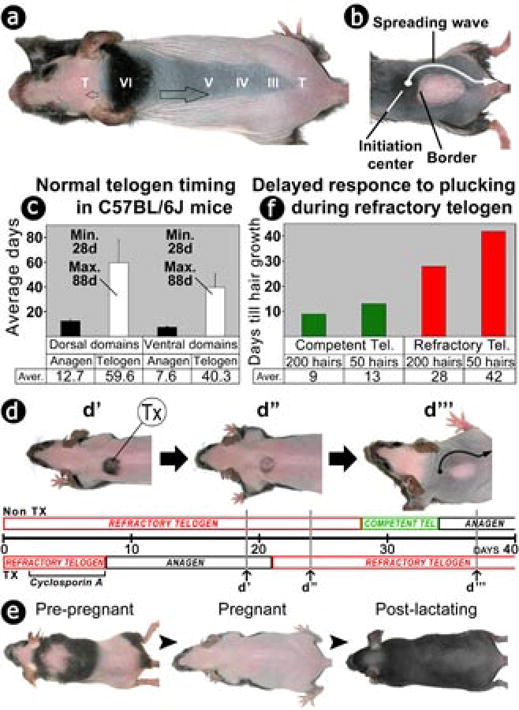
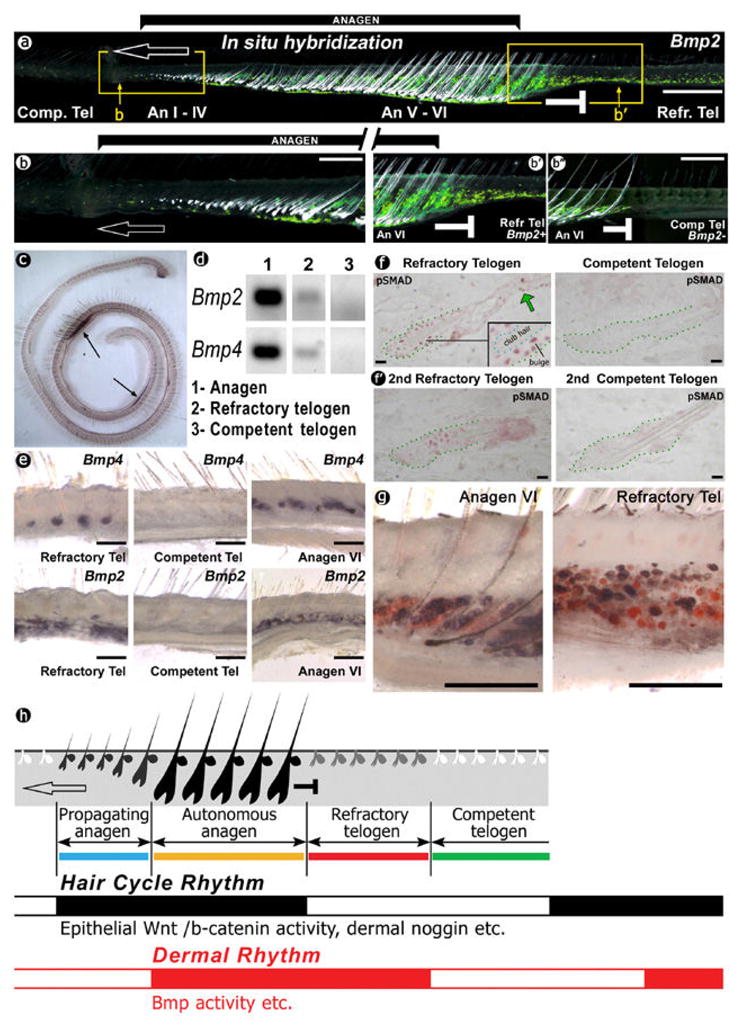
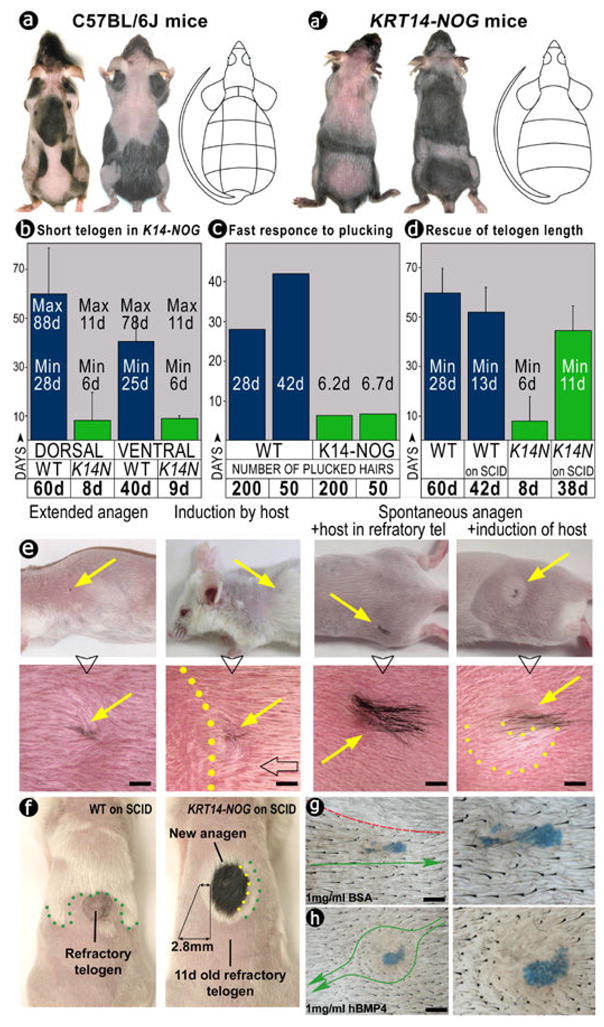
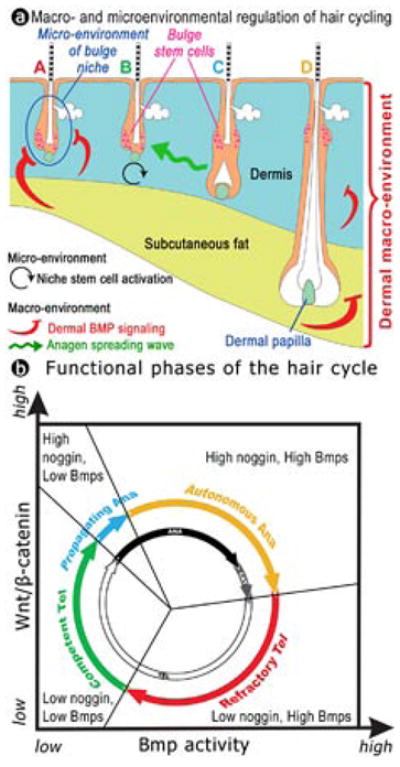
In the age of stem cell engineering it is critical to understand how stem cell activity is regulated during regeneration. Hairs are mini-organs that undergo cyclic regeneration throughout adult life, and are an important model for organ regeneration. Hair stem cells located in the follicle bulge are regulated by the surrounding microenvironment, or niche. The activation of such stem cells is cyclic, involving periodic beta-catenin activity. In the adult mouse, regeneration occurs in waves in a follicle population, implying coordination among adjacent follicles and the extrafollicular environment. Here we show that unexpected periodic expression of bone morphogenetic protein 2 (Bmp2) and Bmp4 in the dermis regulates this process. This BMP cycle is out of phase with the WNT/beta-catenin cycle, thus dividing the conventional telogen into new functional phases: one refractory and the other competent for hair regeneration, characterized by high and low BMP signalling, respectively. Overexpression of noggin, a BMP antagonist, in mouse skin resulted in a markedly shortened refractory phase and faster propagation of the regenerative wave. Transplantation of skin from this mutant onto a wild-type host showed that follicles in donor and host can affect their cycling behaviours mutually, with the outcome depending on the equilibrium of BMP activity in the dermis. Administration of BMP4 protein caused the competent region to become refractory. These results show that BMPs may be the long-sought 'chalone' inhibitors of hair growth postulated by classical experiments. Taken together, results presented in this study provide an example of hierarchical regulation of local organ stem cell homeostasis by the inter-organ macroenvironment. The expression of Bmp2 in subcutaneous adipocytes indicates physiological integration between these two thermo-regulatory organs. Our findings have practical importance for studies using mouse skin as a model for carcinogenesis, intra-cutaneous drug delivery and stem cell engineering studies, because they highlight the acute need to differentiate supportive versus inhibitory regions in the host skin.
Figures
References
-
- Stenn KS, Paus R. Controls of hair follicle cycling. Physiol Rev. 2001;81:449–494. - PubMed
-
- Morris RJ, Liu Y, Marles L, Yang Z, Trempus C, et al. Capturing and profiling adult hair follicle stem cells. Nat Biotechnol. 2004;22:411–7. - PubMed
-
- Fuchs E, Tumbar T, Guasch G. Socializing with the neighbors: stem cells and their niche. Cell. 2004;116:769–78. - PubMed
-
- Huelsken J, Vogel R, Erdmann B, Cotsarelis G, Birchmeier W. beta-Catenin controls hair follicle morphogenesis and stem cell differentiation in the skin. Cell. 2001;105:533–45. - PubMed
-
- Reddy S, Andl T, Bagasra A, Lu MM, Epstein DJ, et al. Characterization of Wnt gene expression in developing and postnatal hair follicles and identification of Wnt5a as a target of Sonic hedgehog in hair follicle morphogenesis. Mech Dev. 2001;107:69–82. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
