

Methionine and its derivatives increase bladder excitability by inhibiting stretch-dependent K(+) channels
- PMID: 18204472
- PMCID: PMC2275456
- DOI: 10.1038/sj.bjp.0707690
Methionine and its derivatives increase bladder excitability by inhibiting stretch-dependent K(+) channels
Abstract
Background and purpose: During the bladder filling phase, the volume of the urinary bladder increases dramatically, with only minimal increases in intravesical pressure. To accomplish this, the smooth muscle of the bladder wall must remain relaxed during bladder filling. However, the mechanisms responsible for the stabilization of bladder excitability during stretch are unclear. We hypothesized that stretch-dependent K(+) (TREK) channels in bladder smooth muscle cells may inhibit contraction in response to stretch.
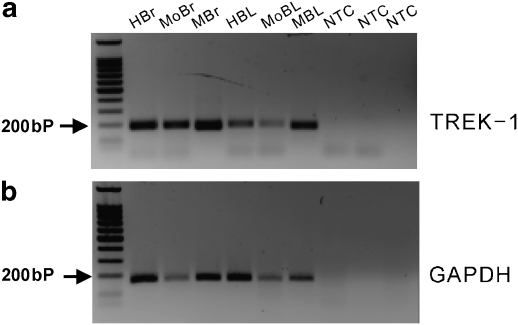
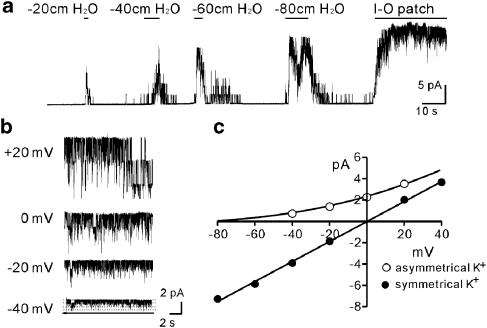
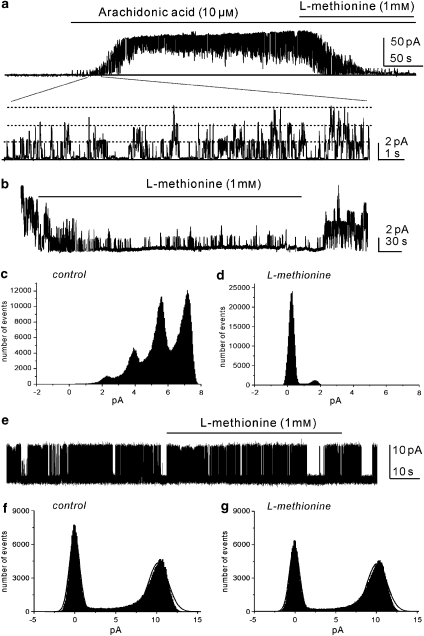
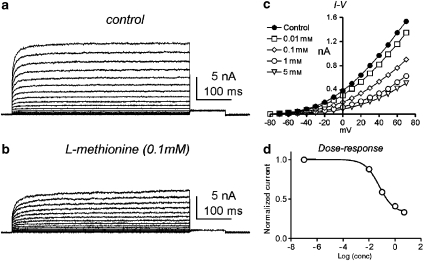
Experimental approaches: Bladder tissues from mouse, guinea pig and monkey were used for molecular, patch clamp, mechanical, electrical, Ca(2+) imaging and cystometric responses to methionine and its derivatives, which are putative blockers of stretch-dependent K(+) (SDK) channels.
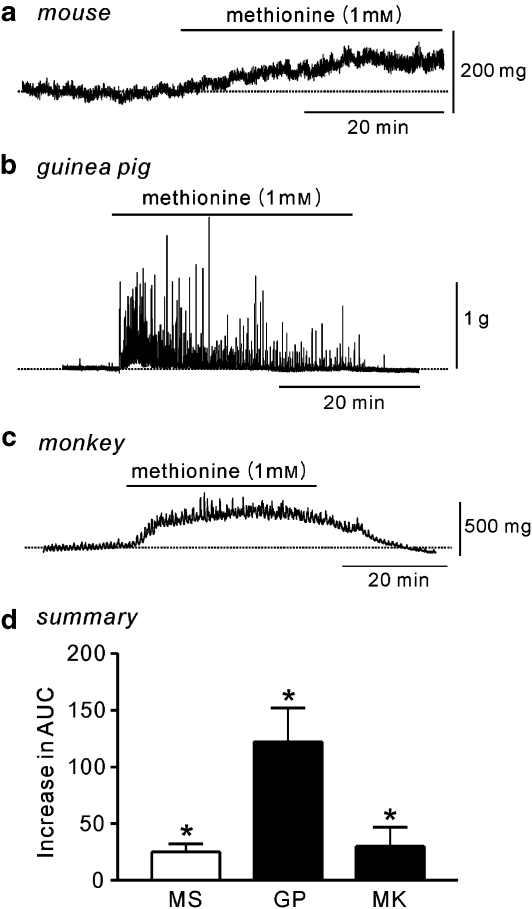
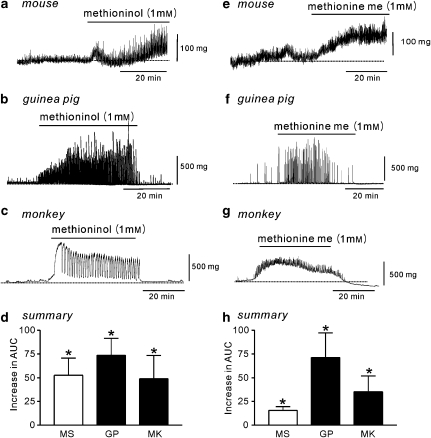
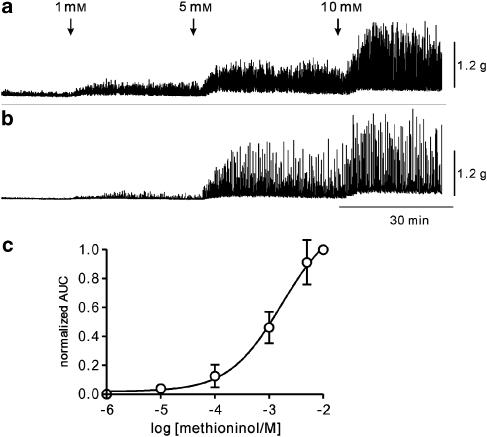
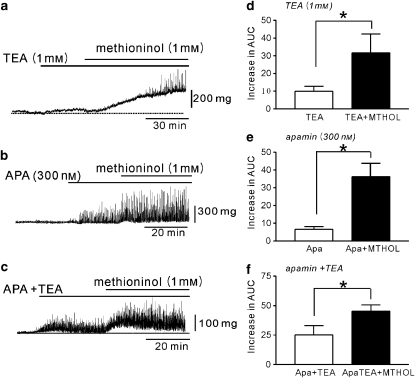
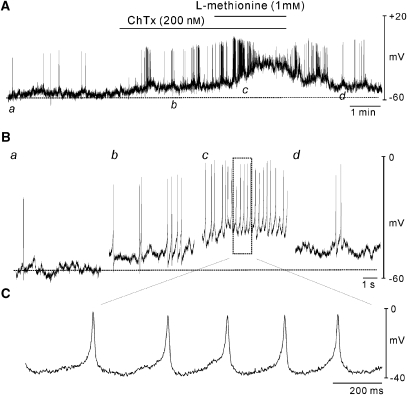
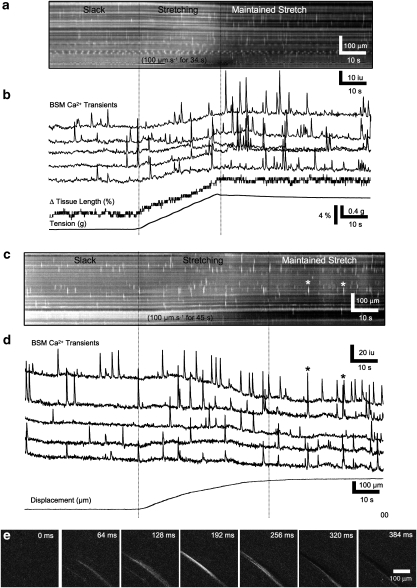
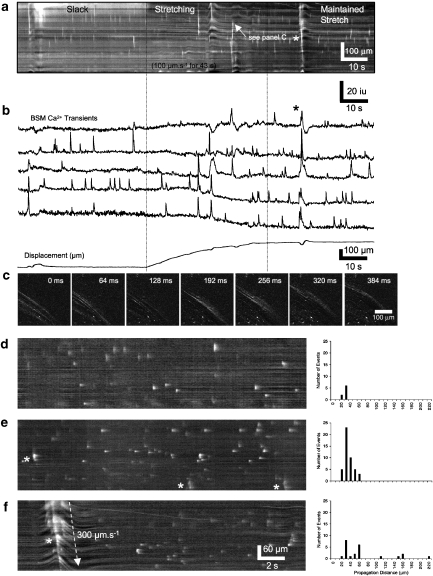
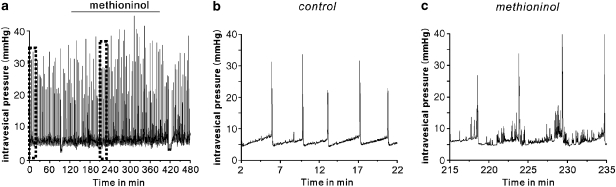
Key results: SDK channels are functionally expressed in bladder myocytes. The single channel conductance of SDK channels is 89pS in symmetrical K(+) conditions and is blocked by L-methionine. Expressed TREK-1 currents are also inhibited by L-methioninol. All three types of bladder smooth muscle cells from mouse, guinea pig and monkey expressed TREK-1 genes. L-methionine, methioninol and methionine methyl ester but not D-methionine increased contractility in concentration-dependent manner. Methioninol further increased contractility and depolarized the membrane in the presence of blockers of Ca(2+)-activated K(+) conductance. L-methionine induced Ca(2+) waves that spread long distances through the tissue under stretched conditions and were associated with strong contractions. In cystometric assays, methioninol injection increased bladder excitability mimicking overactive bladder activity.
Conclusions and implications: Methioninol-sensitive K(+) (SDK, TREK-1) channels appear to be important to prevent spread of excitation through the syncitium during bladder filling.
Figures
References
-
- Bayguinov O, Hagen B, Bonev AD, Nelson MT, Sanders KM. Intracellular calcium events activated by ATP in murine colonic myocytes. Am J Physiol Cell Physiol. 2000;279:C126–C135. - PubMed
-
- Brading AF. Ion channels and control of contractile activity in urinary bladder smooth muscle. Jpn J Pharmacol. 1992;58:120P–127P. - PubMed
-
- Brading AF. The pathophysiological changes in the bladder obstructed by benign prostatic hyperplasia. Br J Urol. 1994;74:133. - PubMed
-
- Dick GM, Bradley KK, Horowitz B, Hume JR, Sanders KM. Functional and molecular identification of a novel chloride conductance in canine colonic smooth muscle. Am J Physiol. 1998;275:C940–C950. - PubMed
-
- Farrugia G, Holm AN, Rich A, Sarr MG, Szurszewski JH, Rae JL. A mechanosensitive calcium channel in human intestinal smooth muscle cells. Gastroenterology. 1999;117:900–905. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
