

Characterization of a juvenile hormone-regulated chymotrypsin-like serine protease gene in Aedes aegypti mosquito
- PMID: 18207080
- PMCID: PMC2253661
- DOI: 10.1016/j.ibmb.2007.10.008
Characterization of a juvenile hormone-regulated chymotrypsin-like serine protease gene in Aedes aegypti mosquito
Abstract
After female mosquitoes ingest blood from vertebrate hosts, exopeptidases and endopeptidases are required for digesting blood proteins in the midgut into amino acids, which female mosquitoes use to build yolk proteins. These proteases are not always present in the midgut, and their diverse expression patterns suggest that production of these enzymes is highly regulated in order to meet specific physiological demands at various stages. Here we report identification of a serine-type protease, JHA15, in the yellow fever mosquito Aedes aegypti. This protein shares high sequence homology with chymotrypsins, and indeed exhibits specific chymotrypsin enzymatic activity. The JHA15 gene is expressed primarily in the midgut of adult female mosquitoes. Our results indicate that its transcription is activated by juvenile hormone in the newly emerged female adults. Although its mRNA profile is similar to that of the early trypsin gene, we found that JHA15 proteins were readily detected in the midgut epithelium cells of both non-blood-fed and blood-fed mosquitoes. Analysis of polysomal RNA further substantiated that synthesis of JHA15 occurs before and shortly after blood feeding. Knocking down expression of JHA15 resulted in no evident phenotypic changes, implying that functional redundancy exists among those proteolytic enzymes.
Figures
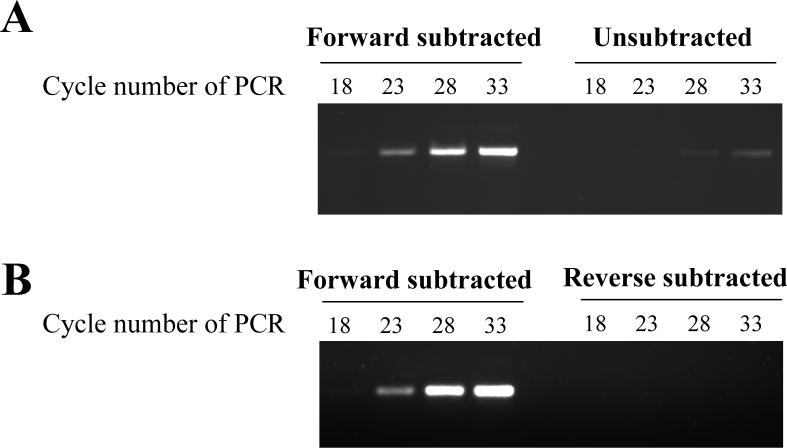

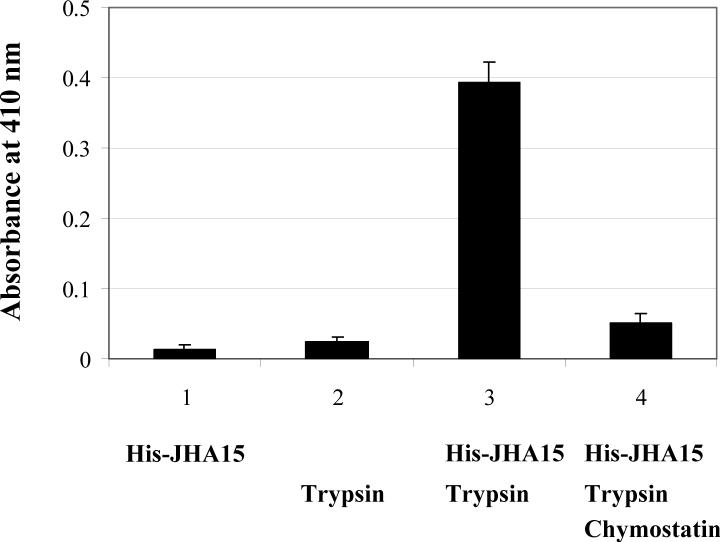
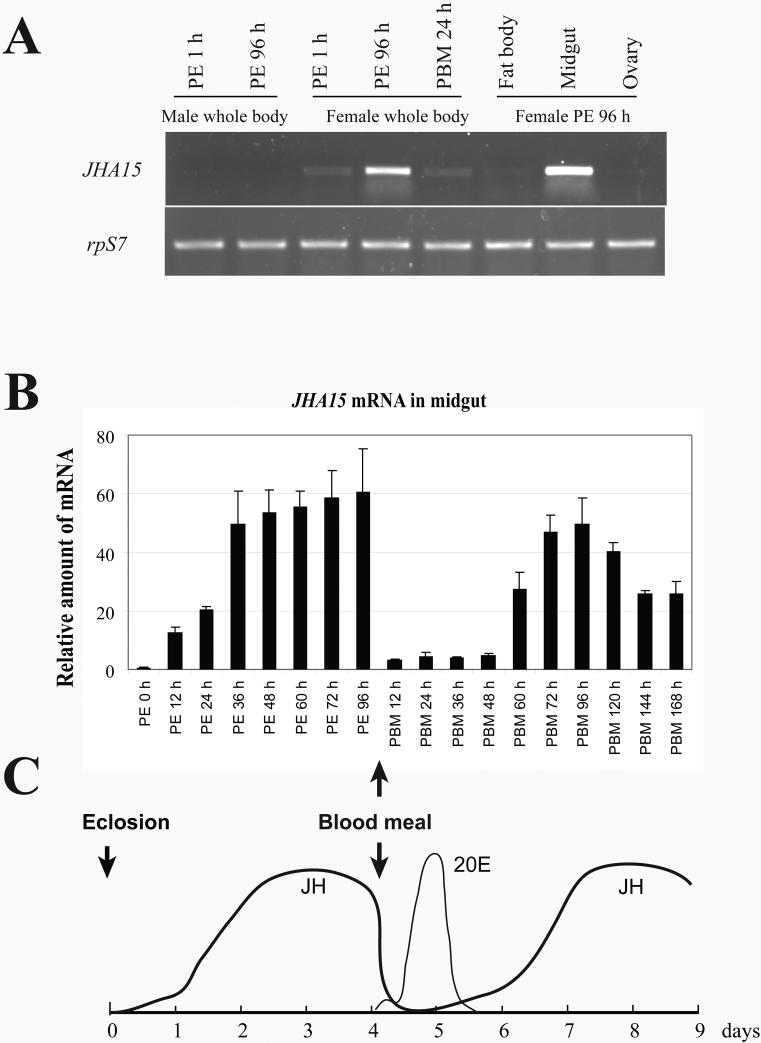


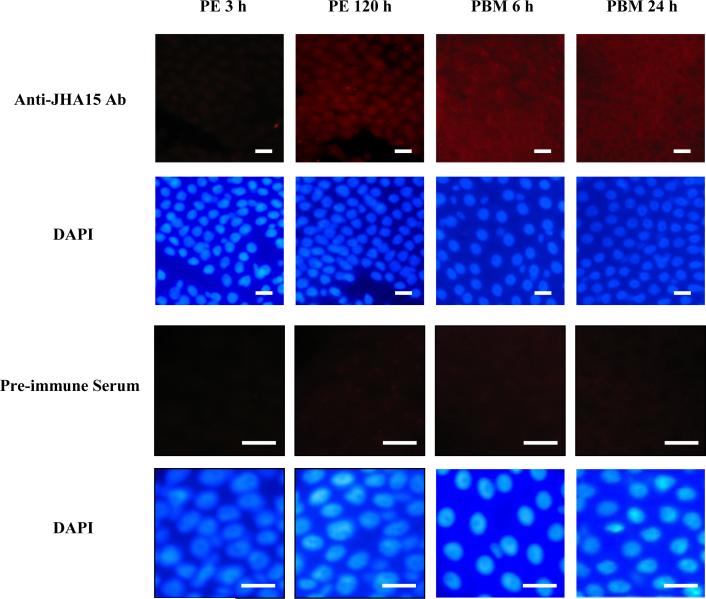


References
-
- Abraham EG, Islam S, Srinivasan P, Ghosh AK, Valenzuela JG, Ribeiro JMC, Kafatos FC, Dimopoulos G, Jacobs-Lorena M. Analysis of the Plasmodium and Anopheles transcriptional repertoire during ookinete development and midgut invasion. J. Biol. Chem. 2004;279:5573–5580. - PubMed
-
- Attardo GM, Hansen IA, Raikhel AS. Nutritional regulation of vitellogenesis in mosquitoes: implications for anautogeny. Insect Biochem. Mol. Biol. 2005;35:661–675. - PubMed
-
- Baker FC, Hagedorn HH, Schooley DA, Wheelock G. Mosquito juvenile hormone: Identification and bioassay activity. J. Insect Physiol. 1983;29:465–470.
-
- Barillas-Mury C, Graf R, Hagedorn HH, Wells MA. cDNA and deduced amino acid sequence of a blood meal-induced trypsin from the mosquito, Aedes aegypti. Insect Biochem. 1991;21:825–831.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
