

Attenuated endocytosis and toxicity of a mutant cholera toxin with decreased ability to cluster ganglioside GM1 molecules
- PMID: 18212085
- PMCID: PMC2292862
- DOI: 10.1128/IAI.01286-07
Attenuated endocytosis and toxicity of a mutant cholera toxin with decreased ability to cluster ganglioside GM1 molecules
Abstract
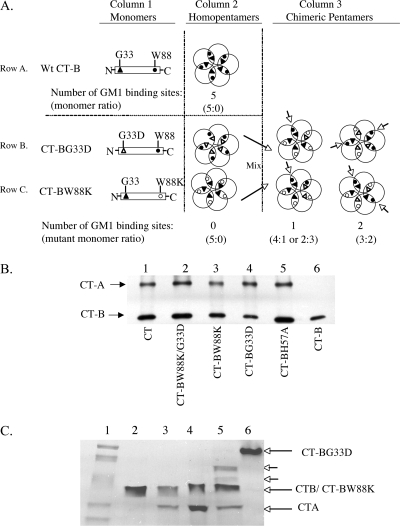
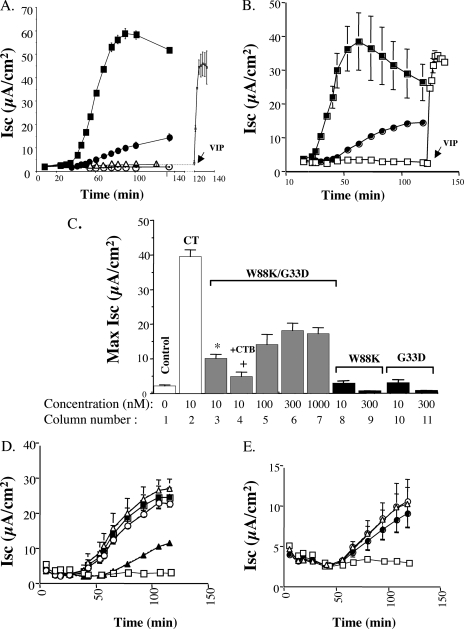
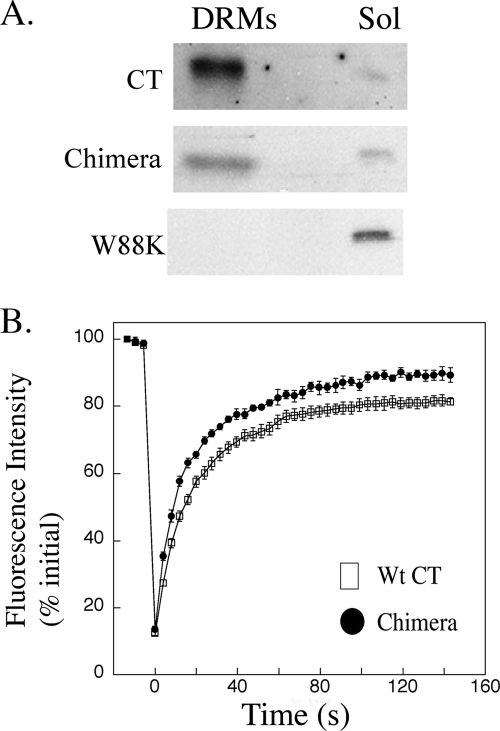
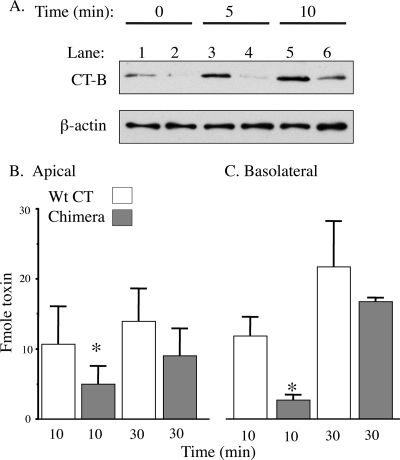
Cholera toxin (CT) moves from the plasma membrane (PM) of host cells to the endoplasmic reticulum (ER) by binding to the lipid raft ganglioside GM(1). The homopentomeric B-subunit of the toxin can bind up to five GM(1) molecules at once. Here, we examined the role of polyvalent binding of GM(1) in CT action by producing chimeric CTs that had B-subunits with only one or two normal binding pockets for GM(1). The chimeric toxins had attenuated affinity for binding to host cell PM, as expected. Nevertheless, like wild-type (wt) CT, the CT chimeras induced toxicity, fractionated with detergent-resistant membranes extracted from toxin-treated cells, displayed restricted diffusion in the plane of the PM in intact cells, and remained bound to GM(1) when they were immunoprecipitated. Thus, binding normally to two or perhaps only one GM(1) molecule is sufficient for association with lipid rafts in the PM and toxin action. The chimeric toxins, however, were much less potent than wt toxin, and they entered the cell by endocytosis more slowly, suggesting that clustering of GM(1) molecules by the B-subunit enhances the efficiency of toxin uptake and perhaps also trafficking to the ER.
Figures
References
-
- Anderson, R. G., and K. Jacobson. 2002. A role for lipid shells in targeting proteins to caveolae, rafts, and other lipid domains. Science 2961821-1825. - PubMed
-
- Brown, D. A., and E. London. 2000. Structure and function of sphingolipid- and cholesterol-rich membrane rafts. J. Biol. Chem. 27517221-17224. - PubMed
-
- Cherukuri, A., T. Shoham, H. W. Sohn, S. Levy, S. Brooks, R. Carter, and S. K. Pierce. 2004. The tetraspanin CD81 is necessary for partitioning of coligated CD19/CD21-B cell antigen receptor complexes into signaling-active lipid rafts. J. Immunol. 172370-380. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
