

Ras nanoclusters: combining digital and analog signaling
- PMID: 18212529
- PMCID: PMC2782570
- DOI: 10.4161/cc.7.2.5237
Ras nanoclusters: combining digital and analog signaling
Abstract
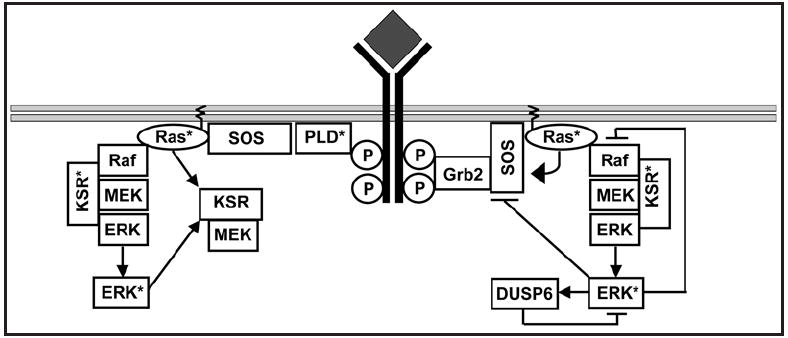
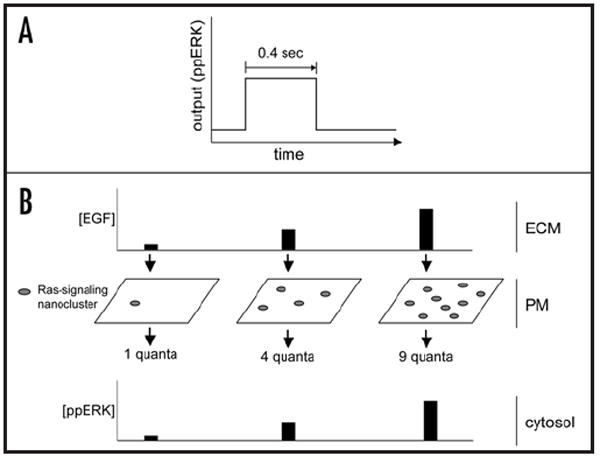
Cellular signaling pathways respond to external inputs to drive pivotal cellular decisions. Far from being mere data relay systems, signaling cascades form complex interacting networks with multiple layers of feedback and feed-forward control loops regulated in both space and time. While it may be intuitively obvious that this complexity allows cells to assess and respond appropriately to a myriad of external cues, untangling the wires to understand precisely how complex networks function as control and computational systems presents a daunting challenge to theoretical and experimental biologists alike. In this review we have focused on activation of the canonical MAP kinase cascade by receptor tyrosine kinases (RTKs) in order to examine some of the fundamental design principles used to build biological circuits and control systems. In particular, we explore how cells can reconfigure signaling cascades to generate distinct biological outputs by utilizing the unique spatial constraints available in biological membranes.
Figures





Similar articles
-
Ras activation of the Raf kinase: tyrosine kinase recruitment of the MAP kinase cascade.Recent Prog Horm Res. 2001;56:127-55. doi: 10.1210/rp.56.1.127. Recent Prog Horm Res. 2001. PMID: 11237210 Review.
-
Plasma membrane nanoswitches generate high-fidelity Ras signal transduction.Nat Cell Biol. 2007 Aug;9(8):905-14. doi: 10.1038/ncb1615. Epub 2007 Jul 8. Nat Cell Biol. 2007. PMID: 17618274
-
Using plasma membrane nanoclusters to build better signaling circuits.Trends Cell Biol. 2008 Aug;18(8):364-71. doi: 10.1016/j.tcb.2008.05.006. Epub 2008 Jul 10. Trends Cell Biol. 2008. PMID: 18620858 Free PMC article. Review.
-
Downregulation of the Ras-mitogen-activated protein kinase pathway by the EphB2 receptor tyrosine kinase is required for ephrin-induced neurite retraction.Mol Cell Biol. 2001 Nov;21(21):7429-41. doi: 10.1128/MCB.21.21.7429-7441.2001. Mol Cell Biol. 2001. PMID: 11585923 Free PMC article.
-
Activation of EphA receptor tyrosine kinase inhibits the Ras/MAPK pathway.Nat Cell Biol. 2001 May;3(5):527-30. doi: 10.1038/35074604. Nat Cell Biol. 2001. PMID: 11331884
Cited by
-
Lessons from computer simulations of Ras proteins in solution and in membrane.Biochim Biophys Acta. 2013 Nov;1830(11):5211-8. doi: 10.1016/j.bbagen.2013.07.024. Epub 2013 Jul 30. Biochim Biophys Acta. 2013. PMID: 23906604 Free PMC article. Review.
-
Tetraspanin CD82 Regulates the Spatiotemporal Dynamics of PKCα in Acute Myeloid Leukemia.Sci Rep. 2016 Jul 15;6:29859. doi: 10.1038/srep29859. Sci Rep. 2016. PMID: 27417454 Free PMC article.
-
N-Ras forms dimers at POPC membranes.Biophys J. 2012 Oct 3;103(7):1585-93. doi: 10.1016/j.bpj.2012.08.043. Epub 2012 Oct 2. Biophys J. 2012. PMID: 23062351 Free PMC article.
-
Akt-ing Up Just About Everywhere: Compartment-Specific Akt Activation and Function in Receptor Tyrosine Kinase Signaling.Front Cell Dev Biol. 2019 May 3;7:70. doi: 10.3389/fcell.2019.00070. eCollection 2019. Front Cell Dev Biol. 2019. PMID: 31131274 Free PMC article. Review.
-
Activation of the MAPK module from different spatial locations generates distinct system outputs.Mol Biol Cell. 2008 Nov;19(11):4776-84. doi: 10.1091/mbc.e08-04-0407. Epub 2008 Sep 10. Mol Biol Cell. 2008. PMID: 18784252 Free PMC article.
References
-
- Schlessinger J. Cell signaling by receptor tyrosine kinases. Cell. 2000;103:211–25. - PubMed
-
- Pawson T. Regulation and targets of receptor tyrosine kinases. Eur J Cancer. 2002;38:3–10. - PubMed
-
- McKay MM, Morrison DK. Integrating signals from RTKs to ERK/MAPK. Oncogene. 2007;26:3113–21. - PubMed
-
- Hancock JF, Magee AI, Childs JE, Marshall CJ. All ras proteins are polyisoprenylated but only some are palmitoylated. Cell. 1989;57:1167–77. - PubMed
-
- Hancock JF, Paterson H, Marshall CJ. A polybasic domain or palmitoylation is required in addition to the CAAX motif to localize p21ras to the plasma membrane. Cell. 1990;63:133–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources