

A two-phase innate host response to alphavirus infection identified by mRNP-tagging in vivo
- PMID: 18215114
- PMCID: PMC2151086
- DOI: 10.1371/journal.ppat.0030199
A two-phase innate host response to alphavirus infection identified by mRNP-tagging in vivo
Abstract
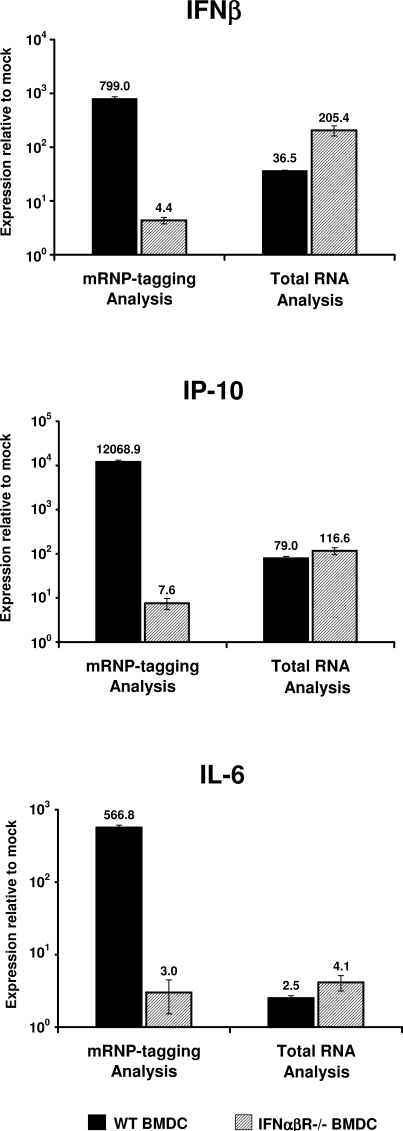
A concept fundamental to viral pathogenesis is that infection induces specific changes within the host cell, within specific tissues, or within the entire animal. These changes are reflected in a cascade of altered transcription patterns evident during infection. However, elucidation of this cascade in vivo has been limited by a general inability to distinguish changes occurring in the minority of infected cells from those in surrounding uninfected cells. To circumvent this inherent limitation of traditional gene expression profiling methods, an innovative mRNP-tagging technique was implemented to isolate host mRNA specifically from infected cells in vitro as well as in vivo following Venezuelan equine encephalitis virus (VEE) infection. This technique facilitated a direct characterization of the host defense response specifically within the first cells infected with VEE, while simultaneous total RNA analysis assessed the collective response of both the infected and uninfected cells. The result was a unique, multifaceted profile of the early response to VEE infection in primary dendritic cells, as well as in the draining lymph node, the initially targeted tissue in the mouse model. A dynamic environment of complex interactions was revealed, and suggested a two-step innate response in which activation of a subset of host genes in infected cells subsequently leads to activation of the surrounding uninfected cells. Our findings suggest that the application of viral mRNP-tagging systems, as introduced here, will facilitate a much more detailed understanding of the highly coordinated host response to infectious agents.
Conflict of interest statement
Figures






Similar articles
-
Replicon particles of Venezuelan equine encephalitis virus as a reductionist murine model for encephalitis.J Virol. 2009 May;83(9):4275-86. doi: 10.1128/JVI.02383-08. Epub 2009 Feb 18. J Virol. 2009. PMID: 19225006 Free PMC article.
-
Early expression of IFN-alpha/beta and iNOS in the brains of Venezuelan equine encephalitis virus-infected mice.J Interferon Cytokine Res. 2000 Feb;20(2):205-15. doi: 10.1089/107999000312621. J Interferon Cytokine Res. 2000. PMID: 10714557
-
Kinetics of cytokine expression and regulation of host protection following infection with molecularly cloned Venezuelan equine encephalitis virus.Virology. 1997 Jul 7;233(2):302-12. doi: 10.1006/viro.1997.8617. Virology. 1997. PMID: 9217054
-
Role of dendritic cell targeting in Venezuelan equine encephalitis virus pathogenesis.J Virol. 2000 Jan;74(2):914-22. doi: 10.1128/jvi.74.2.914-922.2000. J Virol. 2000. PMID: 10623754 Free PMC article.
-
Current Understanding of the Molecular Basis of Venezuelan Equine Encephalitis Virus Pathogenesis and Vaccine Development.Viruses. 2019 Feb 18;11(2):164. doi: 10.3390/v11020164. Viruses. 2019. PMID: 30781656 Free PMC article. Review.
Cited by
-
Comparison of the immune responses induced by chimeric alphavirus-vectored and formalin-inactivated alum-precipitated measles vaccines in mice.PLoS One. 2010 Apr 22;5(4):e10297. doi: 10.1371/journal.pone.0010297. PLoS One. 2010. PMID: 20421972 Free PMC article.
-
Alphavirus replicon particles acting as adjuvants promote CD8+ T cell responses to co-delivered antigen.Vaccine. 2008 Aug 5;26(33):4267-75. doi: 10.1016/j.vaccine.2008.05.046. Epub 2008 Jun 9. Vaccine. 2008. PMID: 18582997 Free PMC article.
-
Infected dendritic cells are sufficient to mediate the adjuvant activity generated by Venezuelan equine encephalitis virus replicon particles.Vaccine. 2012 Jun 22;30(30):4532-42. doi: 10.1016/j.vaccine.2012.04.030. Epub 2012 Apr 21. Vaccine. 2012. PMID: 22531556 Free PMC article.
-
Similarities and differences in antagonism of neuron alpha/beta interferon responses by Venezuelan equine encephalitis and Sindbis alphaviruses.J Virol. 2009 Oct;83(19):10036-47. doi: 10.1128/JVI.01209-09. Epub 2009 Jul 29. J Virol. 2009. PMID: 19641001 Free PMC article.
-
Host Derivation of Sindbis Virus Influences Mammalian Type I Interferon Response to Infection.Viruses. 2023 Aug 3;15(8):1685. doi: 10.3390/v15081685. Viruses. 2023. PMID: 37632027 Free PMC article.
References
-
- Tenenbaum SA, Lager PJ, Carson CC, Keene JD. Ribonomics: identifying mRNA subsets in mRNP complexes using antibodies to RNA-binding proteins and genomic arrays. Methods. 2002;26:191–198. - PubMed
-
- Deo RC, Bonanno JB, Sonenberg N, Burley SK. Recognition of polyadenylate RNA by the poly(A)-binding protein. Cell. 1999;98:835–845. - PubMed
-
- Bernstein P, Ross J. Poly(A), poly(A) binding protein and the regulation of mRNA stability. Trends Biochem Sci. 1989;14:373–377. - PubMed
-
- Burd CG, Dreyfuss G. Conserved structures and diversity of functions of RNA-binding proteins. Science. 1994;265:615–621. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
