

New experiences enhance coordinated neural activity in the hippocampus
- PMID: 18215626
- PMCID: PMC2244590
- DOI: 10.1016/j.neuron.2007.11.035
New experiences enhance coordinated neural activity in the hippocampus
Abstract
The acquisition of new memories for places and events requires synaptic plasticity in the hippocampus, and plasticity depends on temporal coordination among neurons. Spatial activity in the hippocampus is relatively disorganized during the initial exploration of a novel environment, however, and it is unclear how neural activity during the initial stages of learning drives synaptic plasticity. Here we show that pairs of CA1 cells that represent overlapping novel locations are initially more coactive and more precisely coordinated than are cells representing overlapping familiar locations. This increased coordination occurs specifically during brief, high-frequency events (HFEs) in the local field potential that are similar to ripples and is not associated with better coordination of place-specific neural activity outside of HFEs. As novel locations become more familiar, correlations between cell pairs decrease. Thus, hippocampal neural activity during learning has a unique structure that is well suited to induce synaptic plasticity and to allow for rapid storage of new memories.
Figures
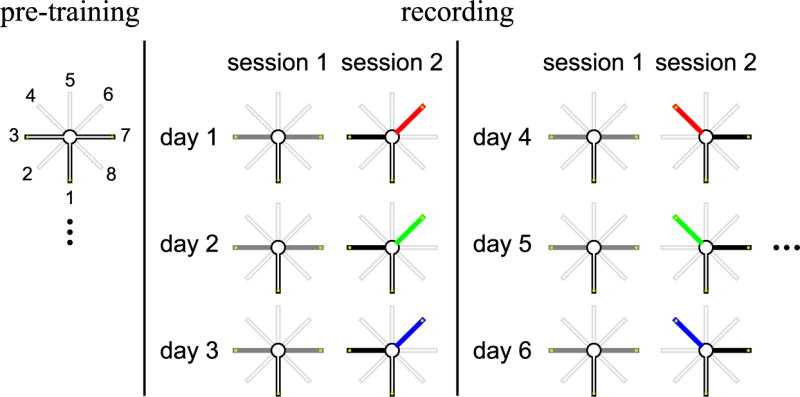
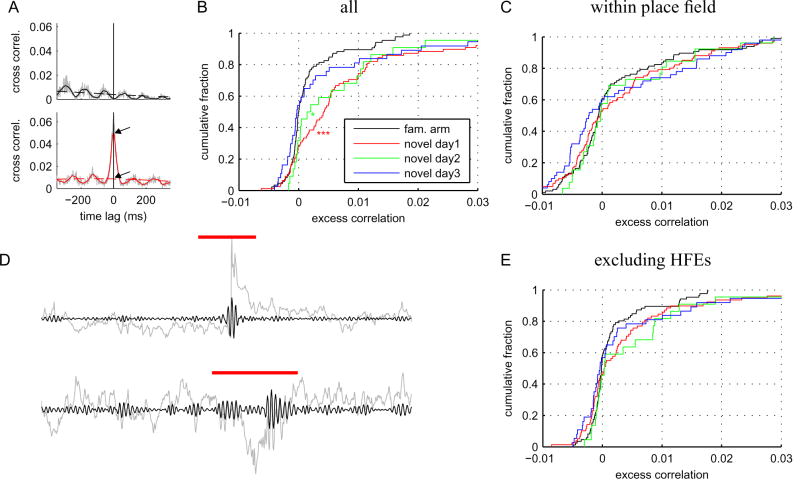
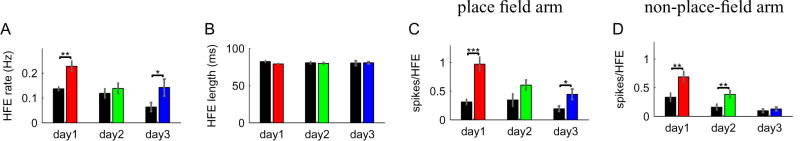
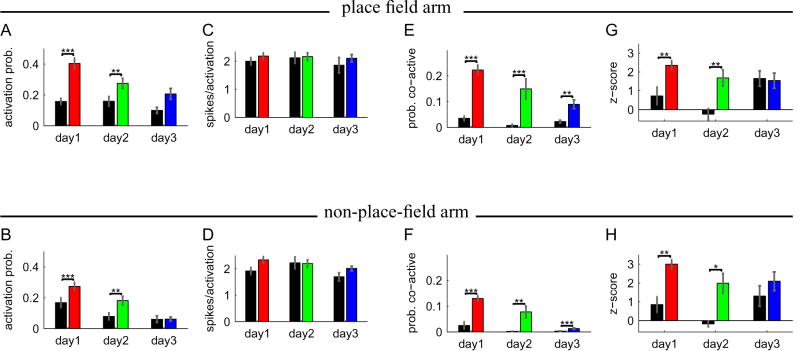
Comment in
-
The ins and outs of hippocampal circuits.Neuron. 2008 Jan 24;57(2):175-7. doi: 10.1016/j.neuron.2008.01.005. Neuron. 2008. PMID: 18215616
Similar articles
-
Selective suppression of hippocampal ripples impairs spatial memory.Nat Neurosci. 2009 Oct;12(10):1222-3. doi: 10.1038/nn.2384. Epub 2009 Sep 13. Nat Neurosci. 2009. PMID: 19749750
-
Temporal encoding of place sequences by hippocampal cell assemblies.Neuron. 2006 Apr 6;50(1):145-57. doi: 10.1016/j.neuron.2006.02.023. Neuron. 2006. PMID: 16600862
-
Fragmentation of grid cell maps in a multicompartment environment.Nat Neurosci. 2009 Oct;12(10):1325-32. doi: 10.1038/nn.2396. Epub 2009 Sep 13. Nat Neurosci. 2009. PMID: 19749749
-
Hippocampal and cortical place cell plasticity: implications for episodic memory.Hippocampus. 2006;16(9):775-84. doi: 10.1002/hipo.20200. Hippocampus. 2006. PMID: 16921502 Review.
-
Parallel processing across neural systems: implications for a multiple memory system hypothesis.Neurobiol Learn Mem. 2004 Nov;82(3):278-98. doi: 10.1016/j.nlm.2004.07.007. Neurobiol Learn Mem. 2004. PMID: 15464410 Review.
Cited by
-
Awake hippocampal sharp-wave ripples support spatial memory.Science. 2012 Jun 15;336(6087):1454-8. doi: 10.1126/science.1217230. Epub 2012 May 3. Science. 2012. PMID: 22555434 Free PMC article.
-
Hippocampal Reactivation Extends for Several Hours Following Novel Experience.J Neurosci. 2019 Jan 30;39(5):866-875. doi: 10.1523/JNEUROSCI.1950-18.2018. Epub 2018 Dec 10. J Neurosci. 2019. PMID: 30530857 Free PMC article.
-
A dentate gyrus-CA3 inhibitory circuit promotes evolution of hippocampal-cortical ensembles during memory consolidation.Elife. 2022 Feb 22;11:e70586. doi: 10.7554/eLife.70586. Elife. 2022. PMID: 35191834 Free PMC article.
-
Neuregulin 1 and ErbB4 Kinase Actively Regulate Sharp Wave Ripples in the Hippocampus.J Neurosci. 2022 Jan 19;42(3):390-404. doi: 10.1523/JNEUROSCI.1022-21.2021. Epub 2021 Nov 29. J Neurosci. 2022. PMID: 34844988 Free PMC article.
-
Supplementation with Achyrocline satureioides Inflorescence Extracts to Pregnant and Breastfeeding Rats Induces Tissue-Specific Changes in Enzymatic Activity and Lower Neonatal Survival.Biomedicines. 2017 Aug 29;5(3):53. doi: 10.3390/biomedicines5030053. Biomedicines. 2017. PMID: 29093434 Free PMC article.
References
-
- Amaral DG, Witter MP. Hippocampal Formation. In: Paxinos C, editor. The Rat Nervous System. Academic Press; 1995. pp. 443–493.
-
- Behrens CJ, van den Boom LP, de HL, Friedman A, Heinemann U. Induction of sharp wave-ripple complexes in vitro and reorganization of hippocampal networks. Nat Neurosci. 2005;8:1560–1567. - PubMed
-
- Brown EN, Barbieri R, Ventura V, Kass RE, Frank LM. The Time-Rescaling Theorem and Its Application to Neural Spike Train Data Analysis. Neural Comput. 2002;14:325–346. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
