

Glucagon-like peptide-1 activation of TCF7L2-dependent Wnt signaling enhances pancreatic beta cell proliferation
- PMID: 18216022
- PMCID: PMC2417166
- DOI: 10.1074/jbc.M706105200
Glucagon-like peptide-1 activation of TCF7L2-dependent Wnt signaling enhances pancreatic beta cell proliferation
Abstract
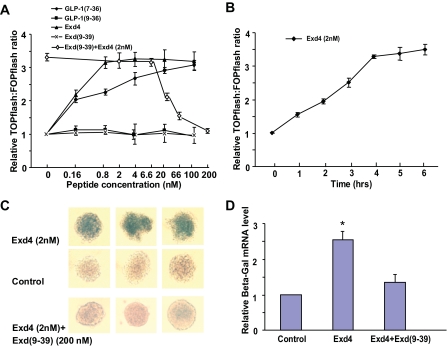
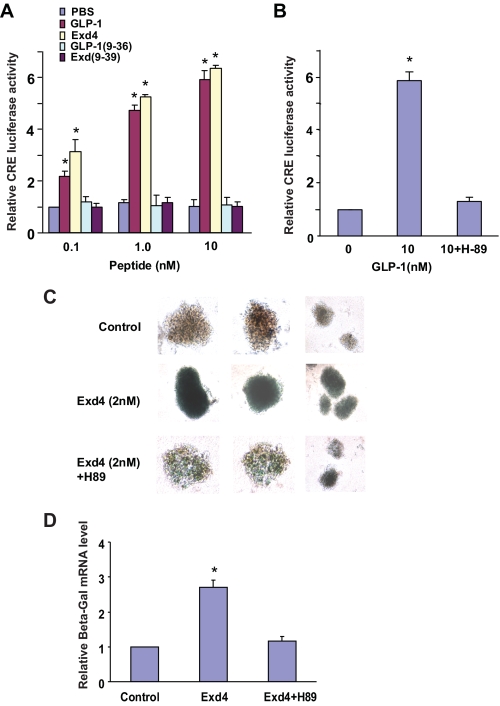
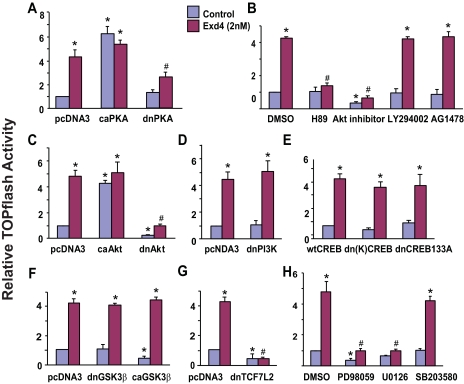
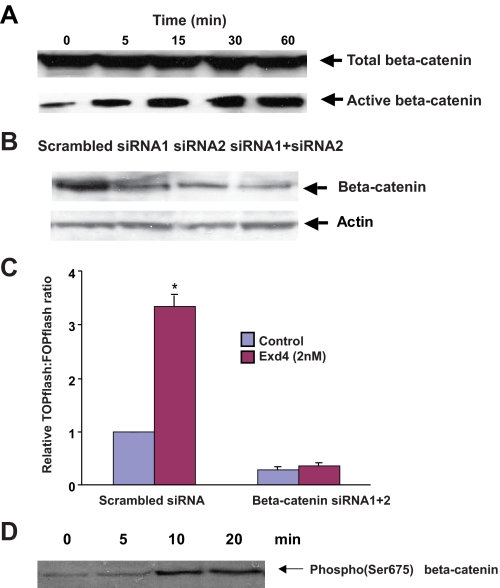
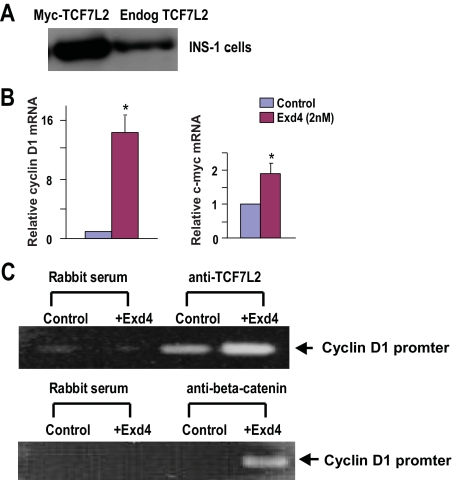
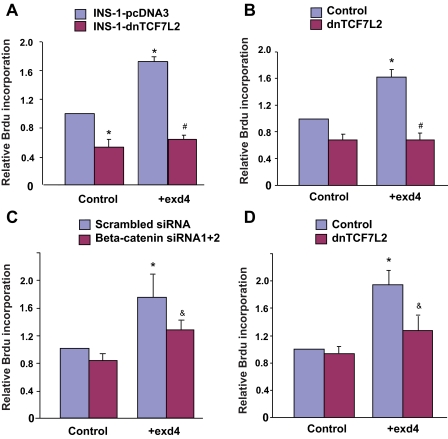
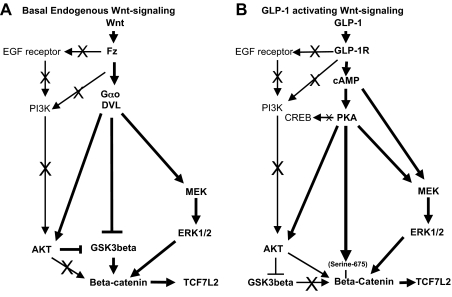
The insulinotropic hormone GLP-1 (glucagon-like peptide-1) is a new therapeutic agent that preserves or restores pancreatic beta cell mass. We report that GLP-1 and its agonist, exendin-4 (Exd4), induce Wnt signaling in pancreatic beta cells, both isolated islets, and in INS-1 cells. Basal and GLP-1 agonist-induced proliferation of beta cells requires active Wnt signaling. Cyclin D1 and c-Myc, determinants of cell proliferation, are up-regulated by Exd4. Basal endogenous Wnt signaling activity depends on Wnt frizzled receptors and the protein kinases Akt and GSK3beta but not cAMP-dependent protein kinase. In contrast, GLP-1 agonists enhance Wnt signaling via GLP-1 receptor-mediated activation of Akt and beta cell independent of GSK3beta. Inhibition of Wnt signaling by small interfering RNAs to beta-catenin or a dominant-negative TCF7L2 decreases both basal and Exd4-induced beta cell proliferation. Wnt signaling appears to mediate GLP-1-induced beta cell proliferation raising possibilities for novel treatments of diabetes.
Figures

References
-
- Holst, J. J. (2004) Expert Opin. Emerg. Drugs 9 155-166 - PubMed
-
- Drucker, D. J., and Nauck, M. A. (2006) Lancet 368 1696-1705 - PubMed
-
- Kieffer, T. J., and Habener, J. F. (1999) Endocr. Rev. 20 876-913 - PubMed
-
- Egan, J. M., Bulotta, A., Hui, H., and Perfetti, R. (2003) Diabetes Metab. Res. Rev. 19 115-123 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
