

Currents in response to rapid concentration jumps of amphetamine uncover novel aspects of human dopamine transporter function
- PMID: 18216205
- PMCID: PMC2459309
- DOI: 10.1523/JNEUROSCI.2796-07.2008
Currents in response to rapid concentration jumps of amphetamine uncover novel aspects of human dopamine transporter function
Abstract
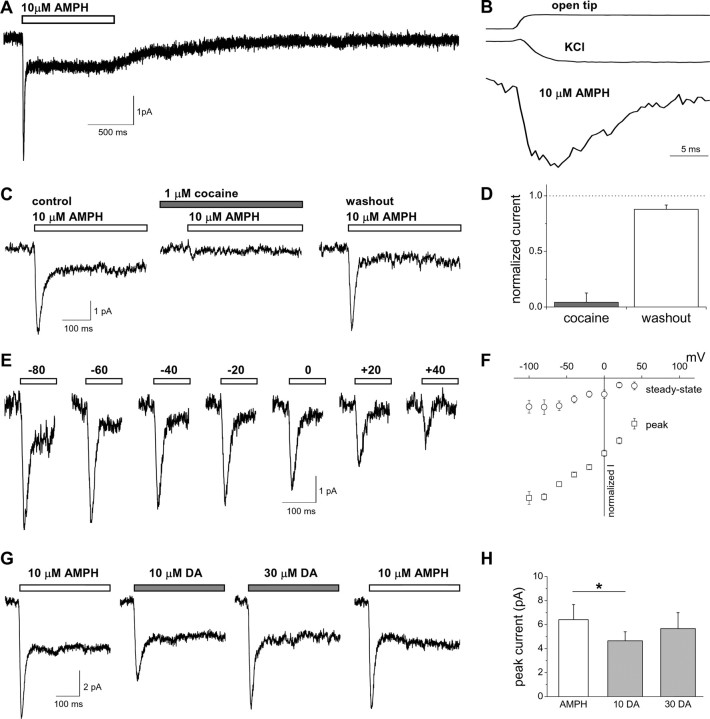
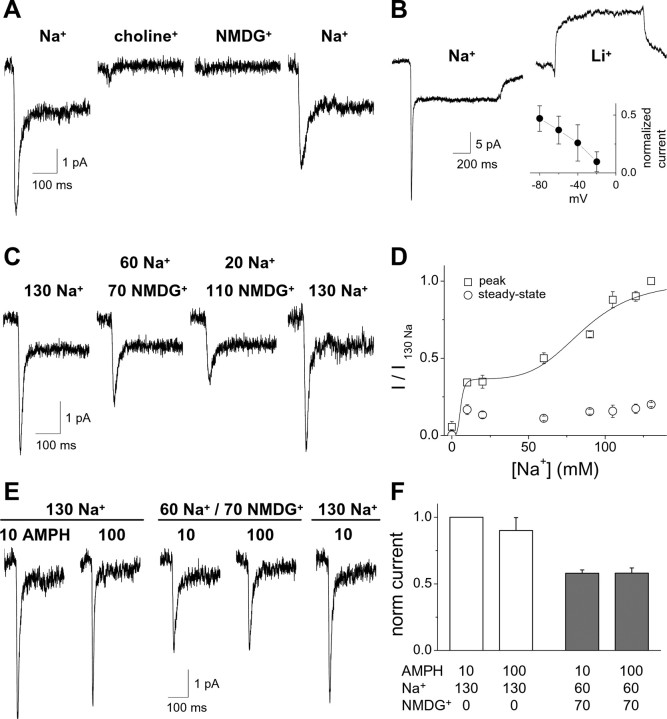
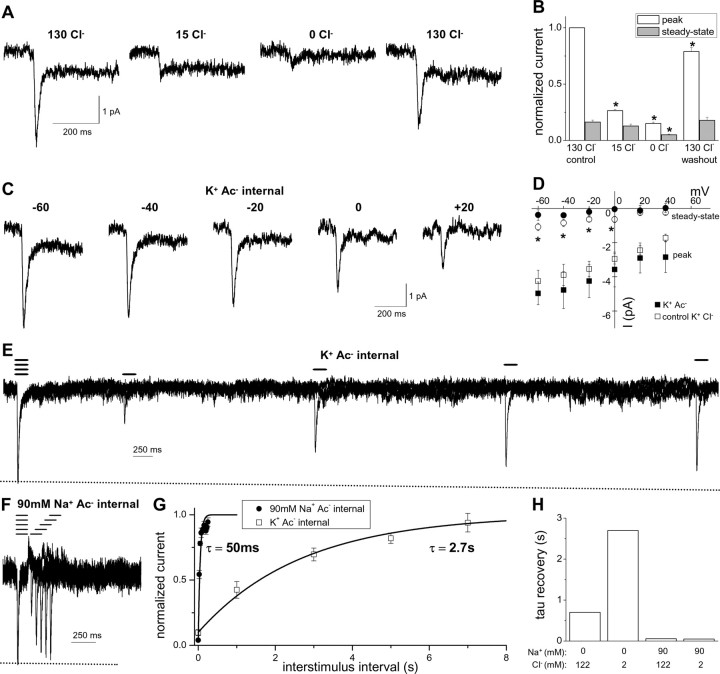
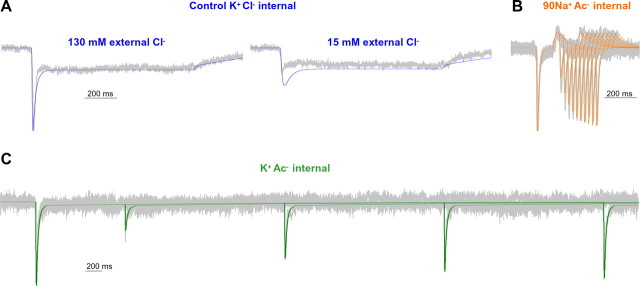
Amphetamine (AMPH) is a widely abused psychostimulant that acts as a substrate for the human dopamine transporter (hDAT). Using a piezoelectric rapid application system, we measured AMPH-induced currents mediated by hDAT. Whole-cell patch-clamp recordings in a heterologous expression system reveal that AMPH induces a rapidly activating and subsequently decaying inward current mediated by hDAT. We hypothesize that this transient inward current reflects a conformational change associated with substrate translocation. The AMPH-induced current strictly depends on extracellular Na+. Elevated intracellular Na+ has no effect on the peak AMPH-induced current amplitude but inhibits the steady-state current. In addition, elevated intracellular Na+ causes an overshoot outward current upon washout of AMPH that reflects hDAT locked in a Na+-exchange mode. Furthermore, elevated intracellular Na+ dramatically accelerates the recovery time from desensitization of the AMPH-induced current, revealing a new role for intracellular Na+ in promoting the transition to the hDAT "outward-facing" conformation. Ion substitution suggests that both extracellular and intracellular Cl- facilitate transporter turnover in contrast to the classical model of Cl- as a cotransported ion. We present an alternating-access model of hDAT function that accurately fits the main features of the experimental data. The model predicts that translocation of substrate occurs within milliseconds of substrate binding but that slow reorientation of the empty transporter is the rate-limiting factor for turnover. The model provides a framework for interpreting perturbations of hDAT activity.
Figures






References
-
- Amara SG, Kuhar MJ. Neurotransmitter transporters: recent progress. Annu Rev Neurosci. 1993;16:73–93. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- DA13975/DA/NIDA NIH HHS/United States
- R01 DA013975/DA/NIDA NIH HHS/United States
- K02 MH057324/MH/NIMH NIH HHS/United States
- NS049335/NS/NINDS NIH HHS/United States
- R01 NS049335/NS/NINDS NIH HHS/United States
- R01 DA011495/DA/NIDA NIH HHS/United States
- K05 DA022413/DA/NIDA NIH HHS/United States
- MH57324/MH/NIMH NIH HHS/United States
- P01 DA012408/DA/NIDA NIH HHS/United States
- R56 NS049335/NS/NINDS NIH HHS/United States
- DA11495/DA/NIDA NIH HHS/United States
- R56 DA013975/DA/NIDA NIH HHS/United States
- DA012408/DA/NIDA NIH HHS/United States
- F32 DA020306/DA/NIDA NIH HHS/United States
LinkOut - more resources
Full Text Sources