

Comparative genomic sequence and expression analyses of Medicago truncatula and alfalfa subspecies falcata COLD-ACCLIMATION-SPECIFIC genes
- PMID: 18218976
- PMCID: PMC2259090
- DOI: 10.1104/pp.107.108779
Comparative genomic sequence and expression analyses of Medicago truncatula and alfalfa subspecies falcata COLD-ACCLIMATION-SPECIFIC genes
Abstract
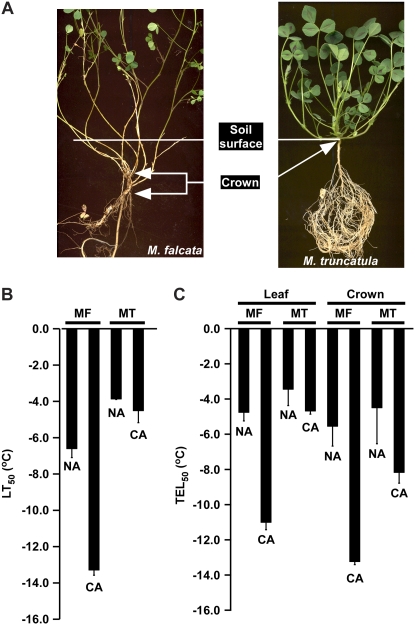
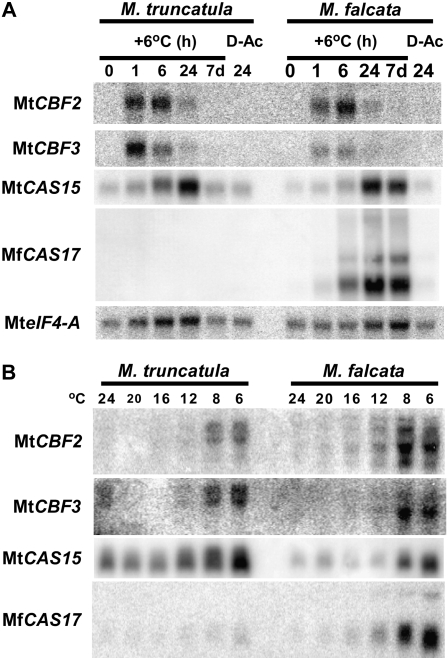
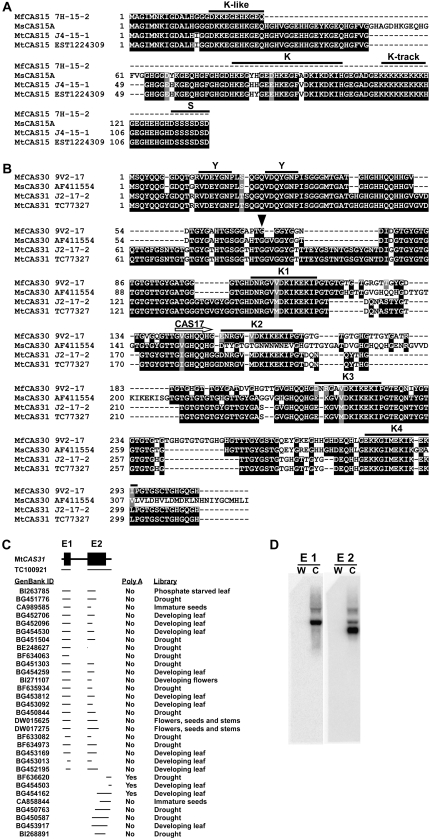
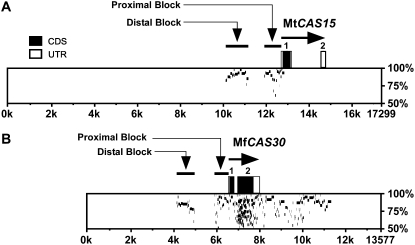
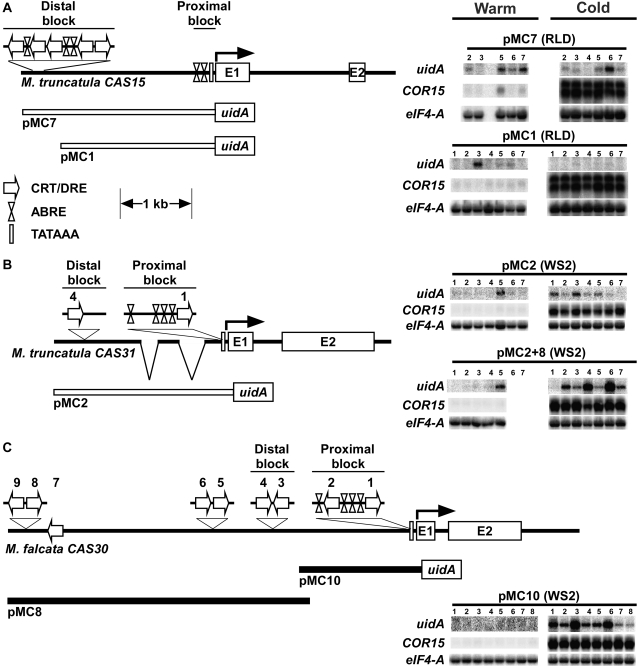
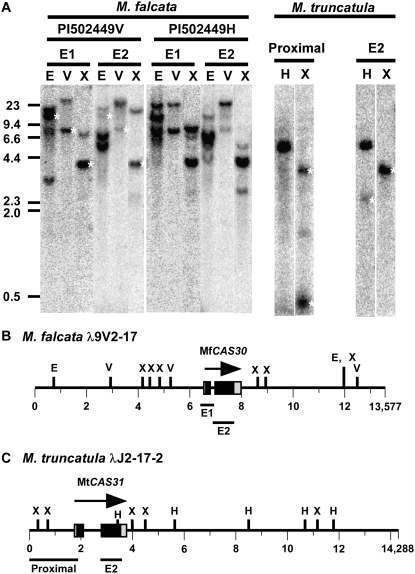
In Arabidopsis (Arabidopsis thaliana) the low-temperature induction of genes encoding the C-REPEAT BINDING FACTOR (CBF) transcriptional activators is a key step in cold acclimation. CBFs in turn activate a battery of downstream genes known as the CBF regulon, which collectively act to increase tolerance to low temperatures. Fundamental questions are: What determines the size and scope of the CBF regulon, and is this is a major determinant of the low-temperature tolerance capacity of individual plant species? Here we have begun to address these questions through comparative analyses of Medicago truncatula and Medicago sativa subsp. falcata. M. truncatula survived to -4 degrees C but did not cold acclimate, whereas Medicago falcata cold acclimated and survived -14 degrees C. Both species possessed low-temperature-induced CBFs but differed in the expression of the COLD-ACCLIMATION-SPECIFIC (CAS) genes, which are candidate CBF targets. M. falcata CAS30 was robustly cold-responsive whereas the MtCAS31 homolog was not. M. falcata also possessed additional CAS30 homologs in comparison to the single CAS31 gene in M. truncatula. MfCAS30 possessed multiple pairs of closely spaced C-REPEAT/DEHYDRATION RESPONSIVE ELEMENT (CRT/DRE) motifs, the cognate CBF binding site in its upstream region whereas MtCAS31 lacked one CRT/DRE partner of the two proximal partner pairs. CAS genes also shared a promoter structure comprising modules proximal and distal to the coding sequence. CAS15, highly cold-responsive in both species, harbored numerous CRT/DRE motifs, but only in the distal module. However, fusion of the MtCAS15 promoter, including the distal module, to a reporter gene did not result in low-temperature responsiveness in stably transformed Arabidopsis. In contrast, both MtCAS31 and MfCAS30 promoter fusions were low-temperature responsive, although the MfCAS31 fusion was less robust than the MfCAS30 fusion. From these studies we conclude that CAS genes harbor CRT/DRE motifs, their proximity to one another is likely key to regulatory output in Medicago, and they may be located kilobases distal to the transcriptional start site. We hypothesize that these differences in CRT/DRE copy numbers in CAS30/CAS31 upstream regions combined with differences in gene copy numbers may be a factor in determining differences in low-temperature tolerance between M. truncatula and M. falcata.
Figures
References
-
- Adams MD (2005) Conserved sequences and the evolution of gene regulatory signals. Curr Opin Genet Dev 15 628–633 - PubMed
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidmen JG, Smith JA, Struhl K (1993) Current Protocols in Molecular Biology. Greene Publishing Associates/John Wiley and Sons, New York
-
- Baker SS, Wilhelm KS, Thomashow MF (1994) The 5′-region of Arabidopsis thaliana cor15a has cis-acting elements that confer cold-, drought- and ABA-regulated gene expression. Plant Mol Biol 24 701–713 - PubMed
-
- Barker DG, Bianchi S, Blondon F, Dattée Y, Duc G, Essad S, Flament P, Gallusci P, Génier G, Guy P, et al (1990) Medicago truncatula, a model plant for studying the molecular genetics of the Rhizobium-legume symbiosis. Plant Mol Biol Rep 8 40–49
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
