

Erythropoietin deficiency decreases vascular stability in mice
- PMID: 18219389
- PMCID: PMC2213374
- DOI: 10.1172/JCI33813
Erythropoietin deficiency decreases vascular stability in mice
Abstract
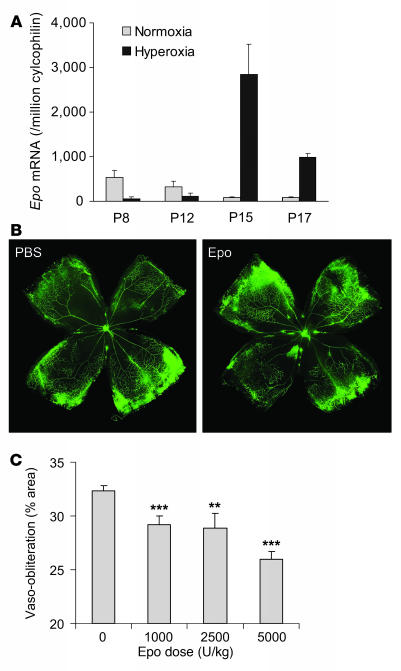
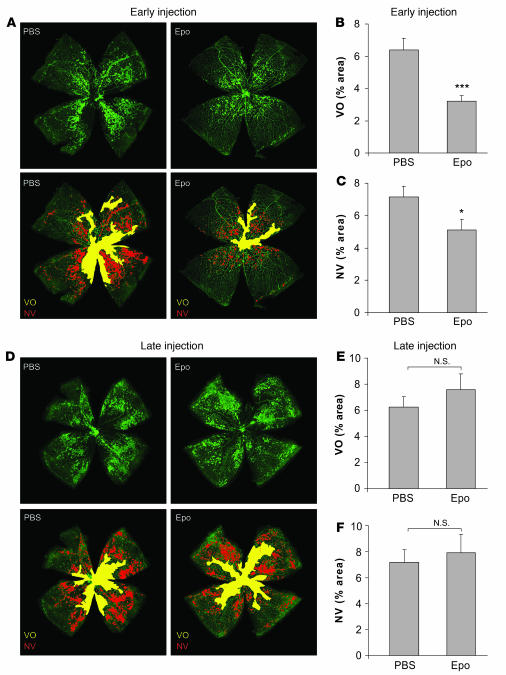
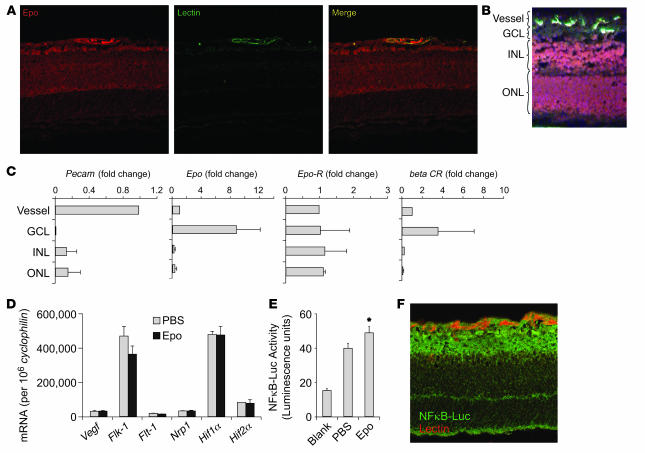
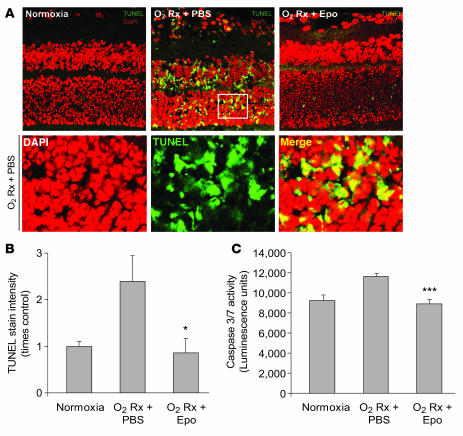
Erythropoietin (Epo), a hormone known to stimulate bone marrow erythrocyte production, is widely used to treat anemia in patients at risk for vascular disease. However, the effects of Epo on angiogenesis are not well defined. We studied the role of Epo in a mouse model of retinopathy characterized by oxygen-induced vascular loss followed by hypoxia-induced pathological neovascularization. Without treatment, local retinal Epo levels were suppressed during the vessel loss phase. Administration of exogenous Epo prevented both vessel dropout and subsequent hypoxia-induced neovascularization. Early use of Epo also protected against hypoxia-induced retinal neuron apoptosis. In contrast, retinal Epo mRNA levels were highly elevated during the retinopathy neovascular phase. Exogenous late Epo treatment did not protect the retina, but rather enhanced pathological neovascularization. Epo's early protective effect occurred through both systemic retinal recruitment of proangiogenic bone marrow-derived progenitor cells and activation of prosurvival NF-kappaB via Epo receptor activation on retinal vessels and neurons. Thus early retinal Epo suppression contributed to retinal vascular instability, and elevated Epo levels during the proliferation stage contributed to neovascularization and disease. Understanding the role of Epo in angiogenesis is critical to timing its intervention in patients with retinopathy or other diseases in which pathological angiogenesis plays a significant role.
Figures
Comment in
-
Erythropoietin: when liability becomes asset in neurovascular repair.J Clin Invest. 2008 Feb;118(2):467-70. doi: 10.1172/JCI34643. J Clin Invest. 2008. PMID: 18219388 Free PMC article.
References
-
- Gariano R.F., Gardner T.W. Retinal angiogenesis in development and disease. Nature. 2005;438:960–966. - PubMed
-
- Sasaki R. Pleiotropic functions of erythropoietin. Intern. Med. 2003;42:142–149. - PubMed
-
- Katsura Y., et al. Erythropoietin is highly elevated in vitreous fluid of patients with proliferative diabetic retinopathy. Diabetes Care. 2005;28:2252–2254. - PubMed
-
- Watanabe D., et al. Erythropoietin as a retinal angiogenic factor in proliferative diabetic retinopathy. N. Engl. J. Med. 2005;353:782–792. - PubMed
-
- Singh A.K., et al. Correction of anemia with epoetin alfa in chronic kidney disease. N. Engl. J. Med. 2006;355:2085–2098. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
