

T cell subset-specific susceptibility to aging
- PMID: 18222733
- PMCID: PMC2435295
- DOI: 10.1016/j.clim.2007.12.002
T cell subset-specific susceptibility to aging
Abstract
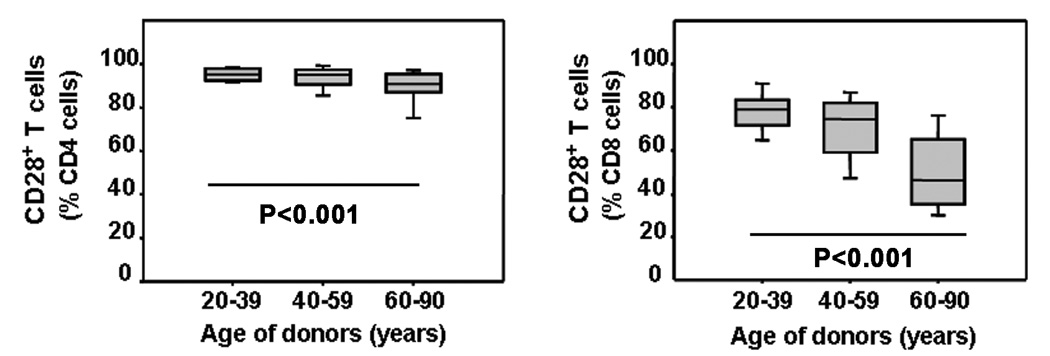
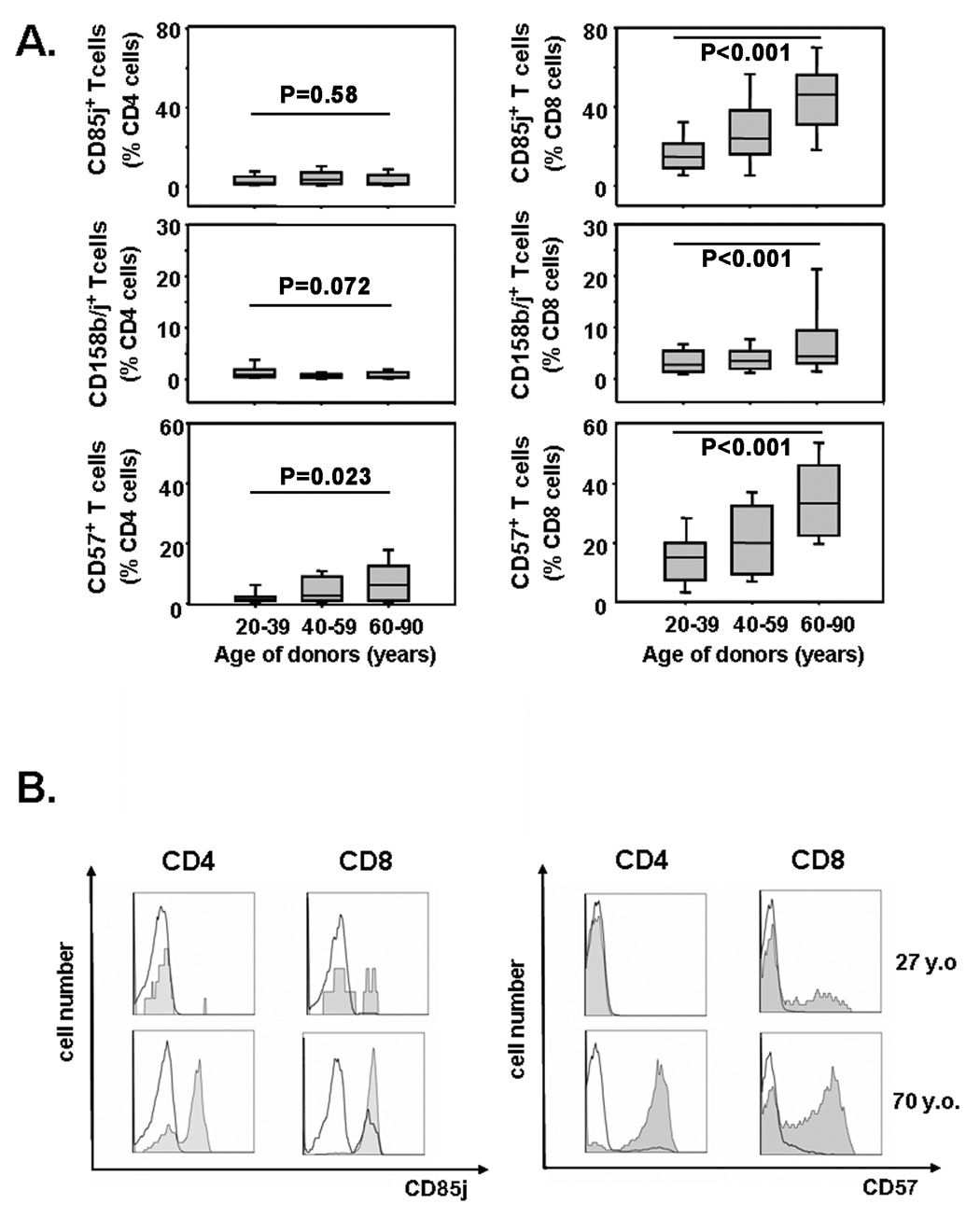
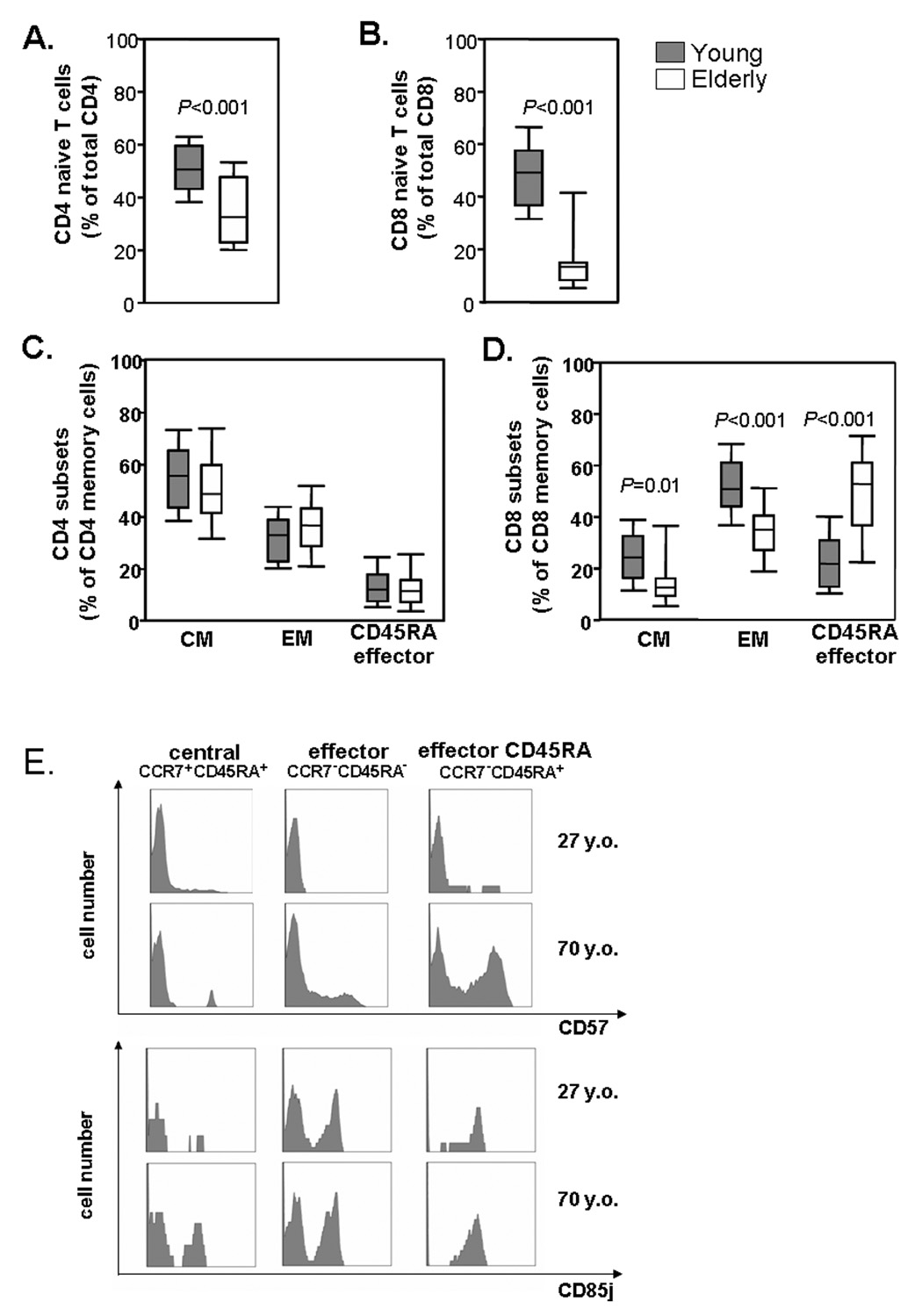
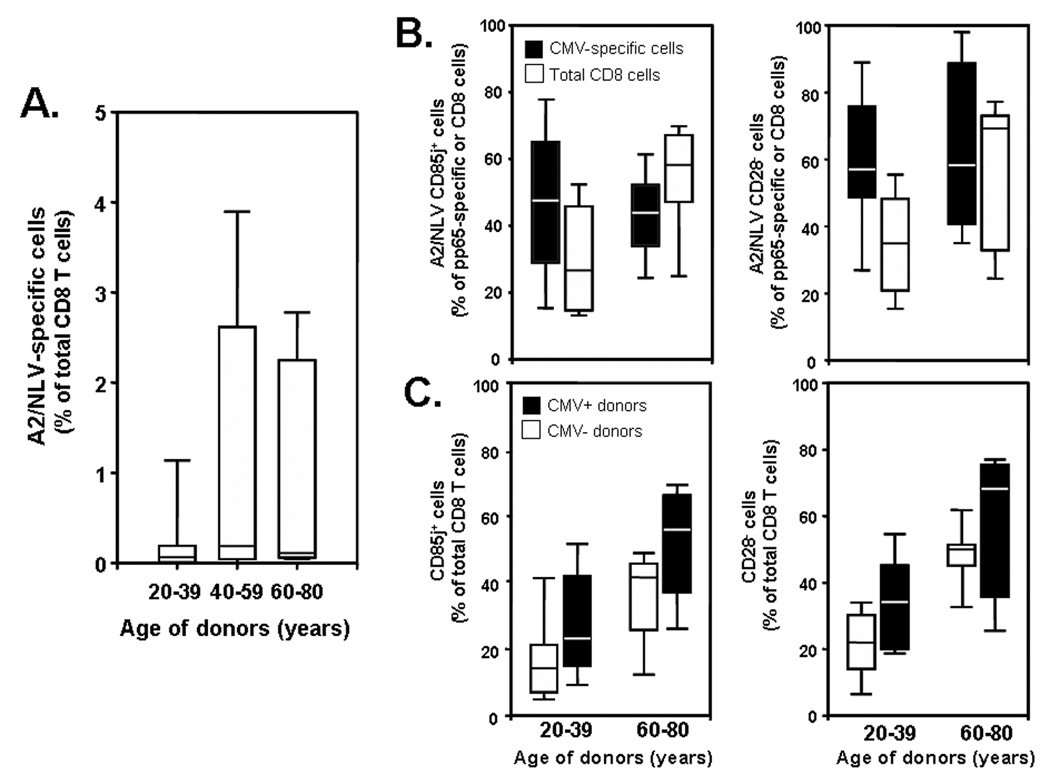
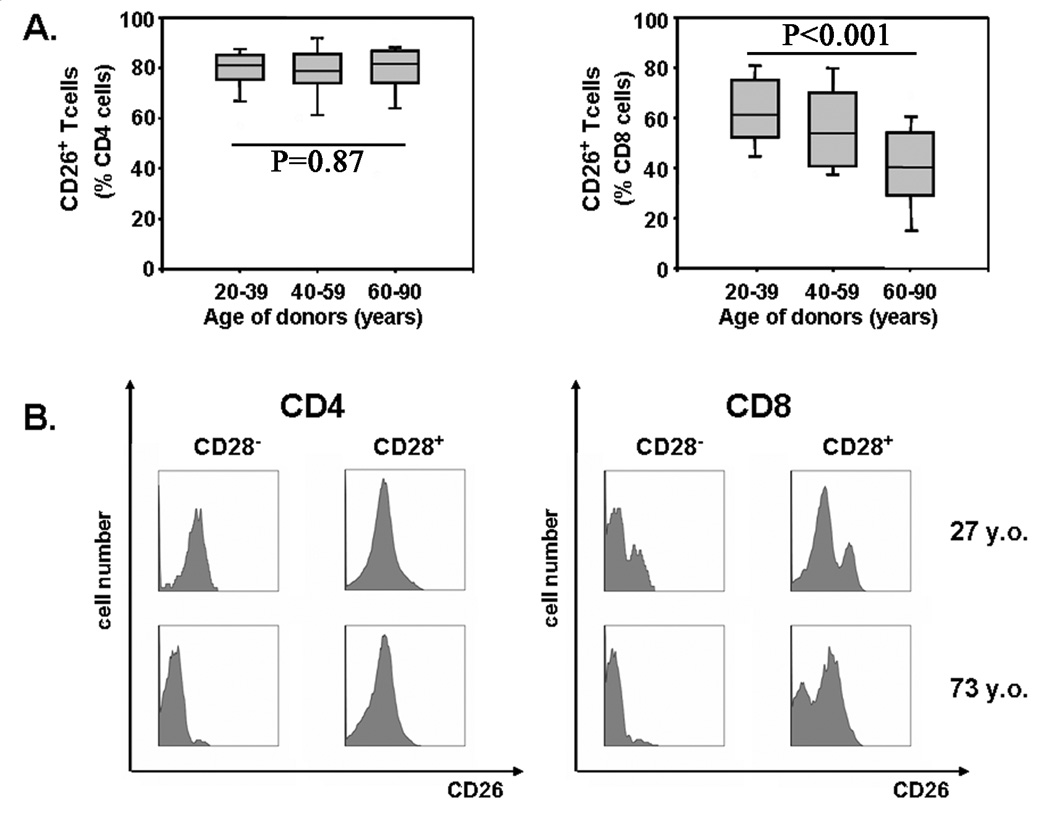
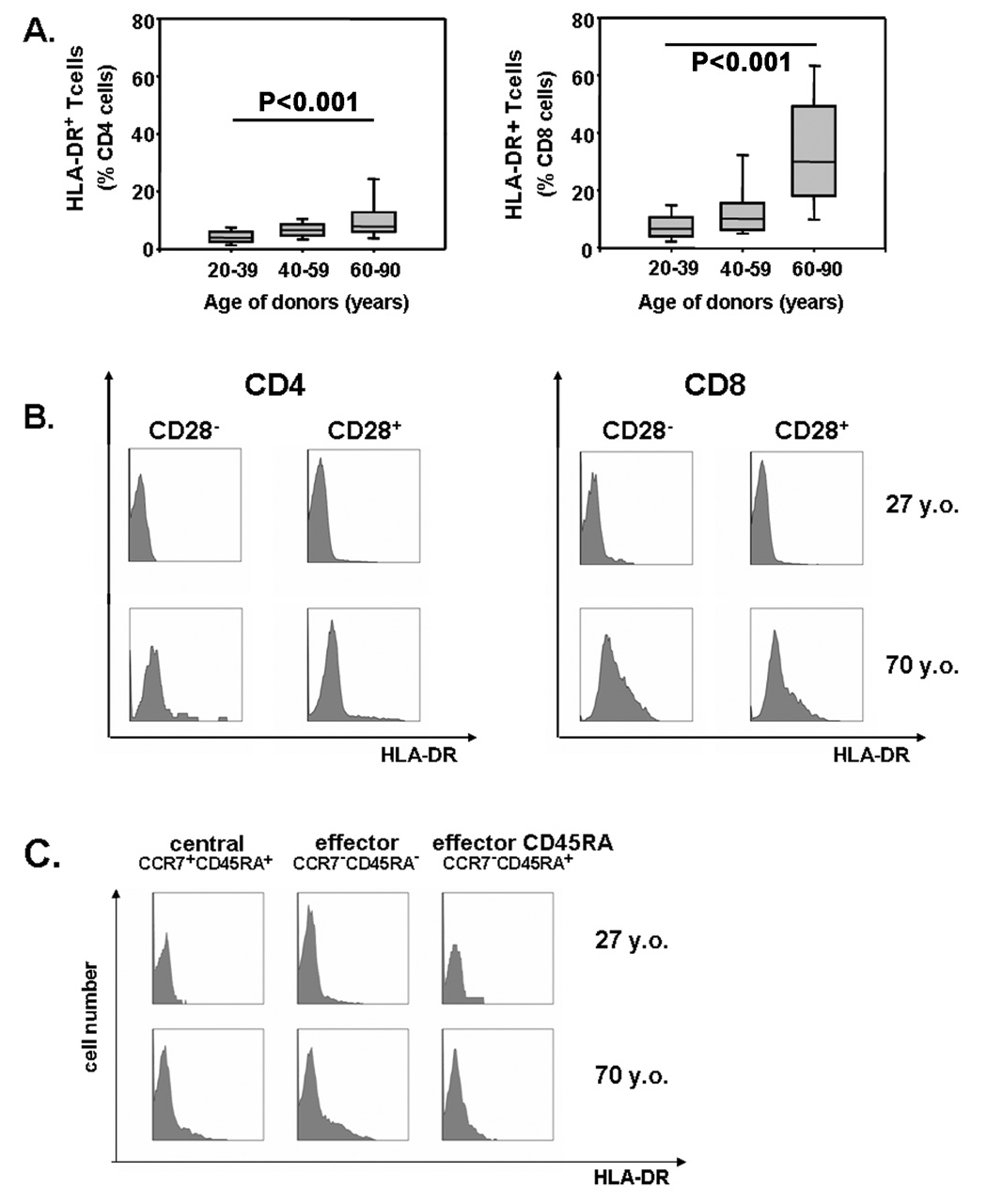
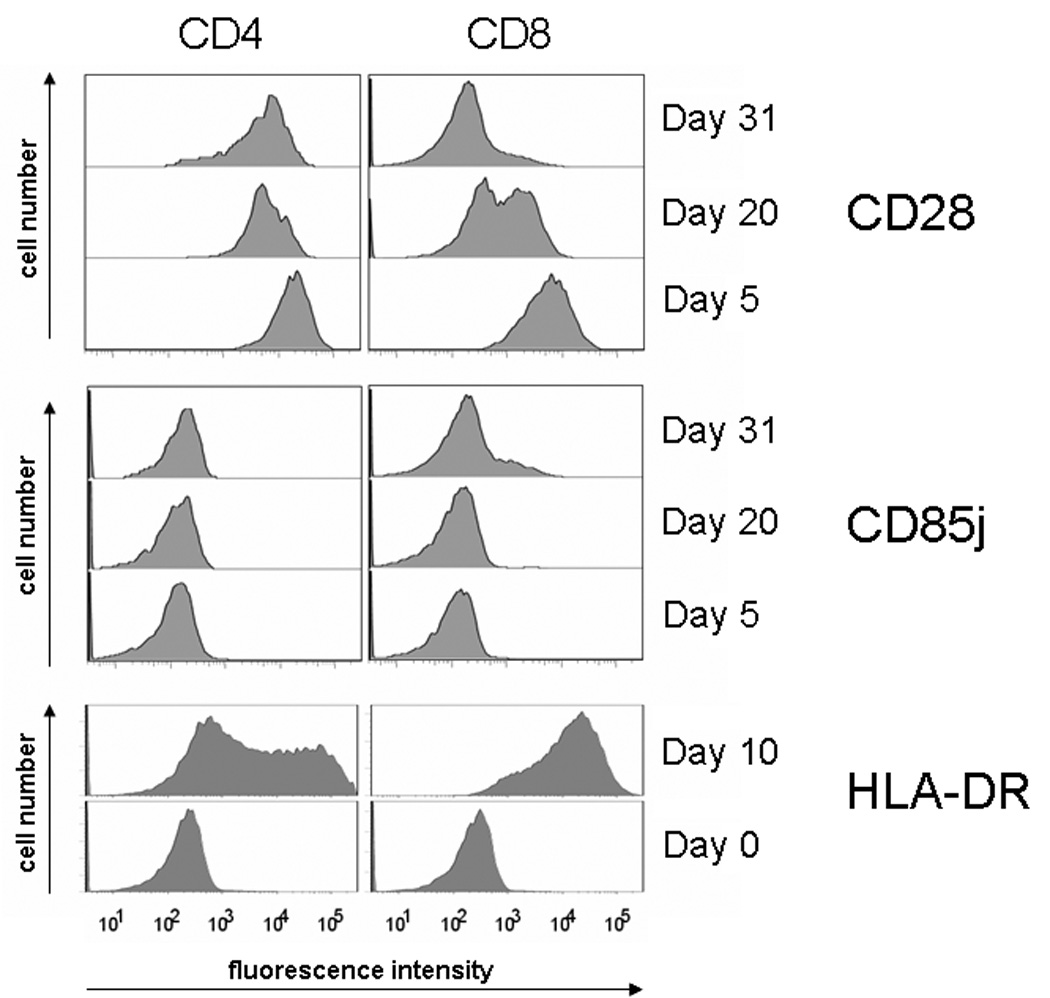
With increasing age, the competence of the immune system to fight infections and tumors declines. Age-dependent changes have been mostly described for human CD8 T cells, raising the question of whether the response patterns for CD4 T cells are different. Gene expression arrays of memory CD4 T cells yielded a similar age-induced fingerprint as has been described for CD8 T cells. In cross-sectional studies, the phenotypic changes were not qualitatively different for CD4 and CD8 T cells, but occurred much more frequently in CD8 T cells. Homeostatic stability partially explained this lesser age sensitivity of CD4 T cells. With aging, naïve and central memory CD8 T cells were lost at the expense of phenotypically distinct CD8 effector T cells, while effector CD4 T cells did not accumulate. However, phenotypic shifts on central memory T cells were also more pronounced in CD8 T cells. This distinct stability in cell surface marker expression can be reproduced in vitro. The data show that CD8 T cells are age sensitive by at least two partially independent mechanisms: fragile homeostatic control and gene expression instability in a large set of regulatory cell surface molecules.
Figures
References
-
- Grubeck-Loebenstein B, Wick G. The aging of the immune system. Adv Immunol. 2002;80:243–284. - PubMed
-
- Goldrath AW, Bevan MJ. Selecting and maintaining a diverse T-cell repertoire. Nature. 1999;402:255–262. - PubMed
-
- Hellerstein MK. Measurement of T-cell kinetics: recent methodologic advances. Immunol Today. 1999;20:438–441. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
