

Thermodynamic and kinetic properties of amino-terminal and S4-S5 loop HERG channel mutants under steady-state conditions
- PMID: 18222997
- PMCID: PMC2367168
- DOI: 10.1529/biophysj.107.116731
Thermodynamic and kinetic properties of amino-terminal and S4-S5 loop HERG channel mutants under steady-state conditions
Abstract
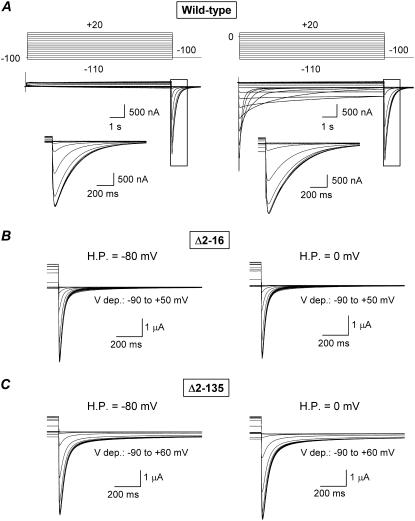
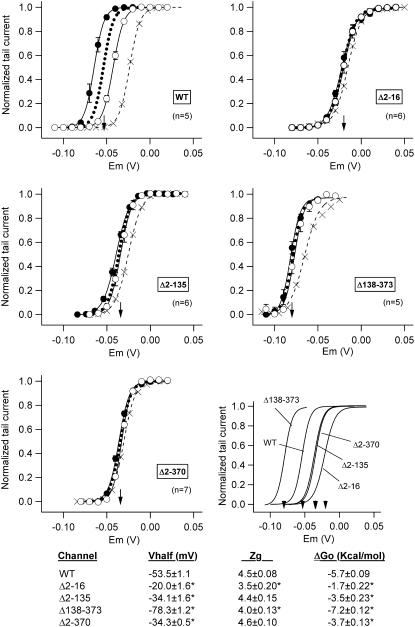
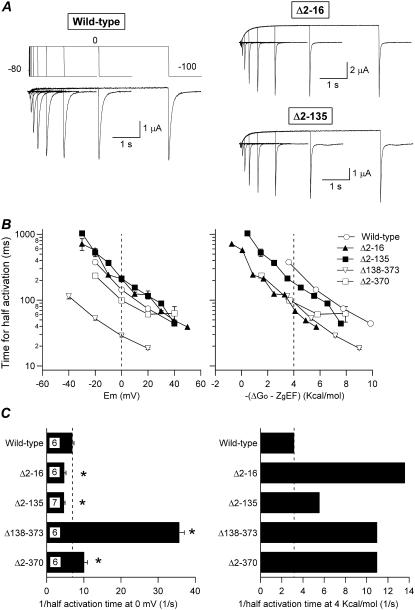
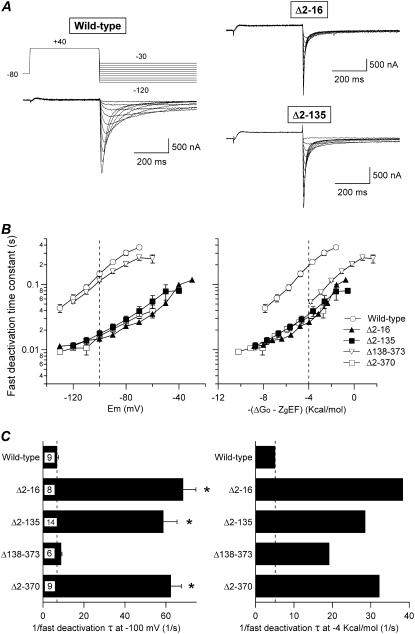
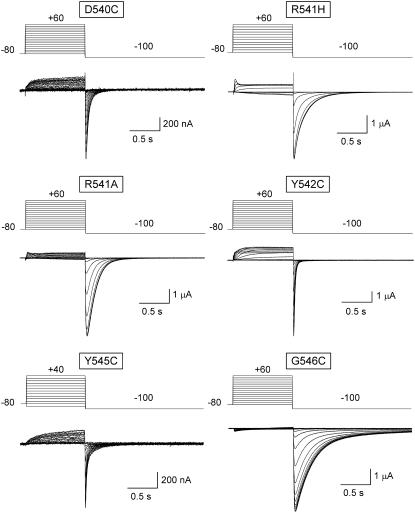
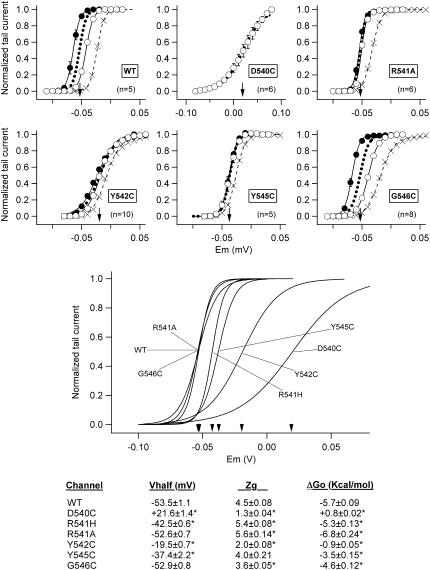
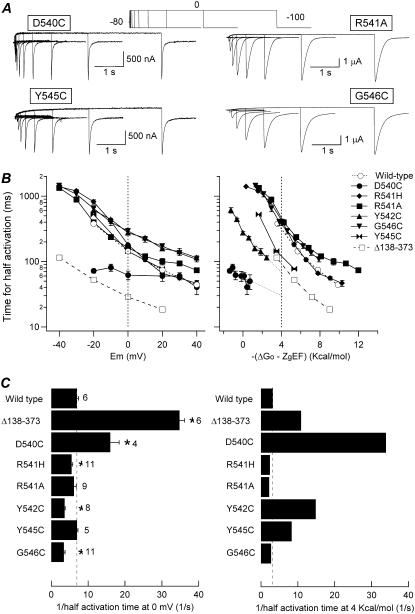
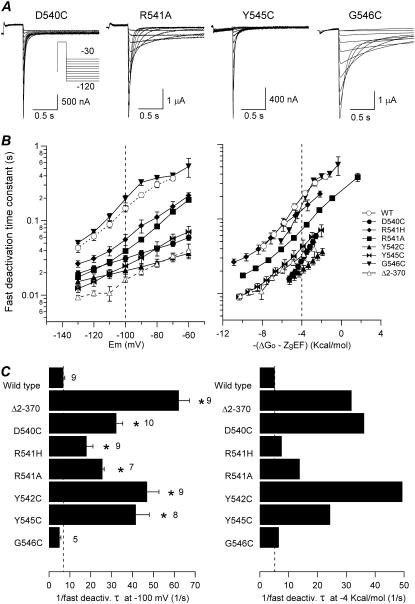
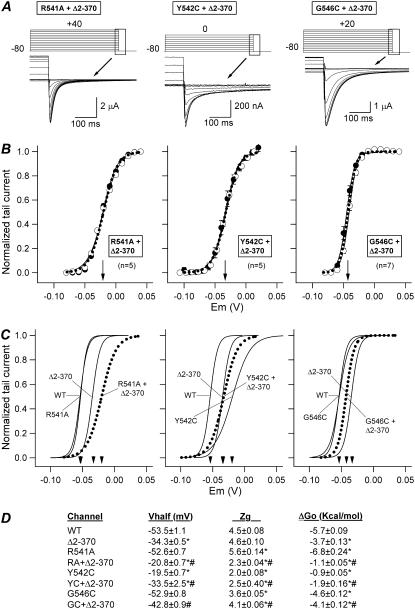
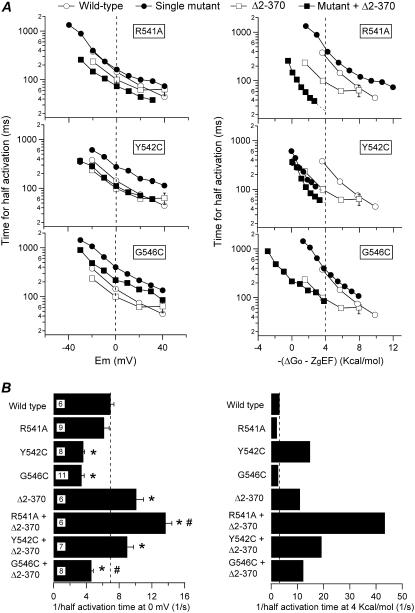
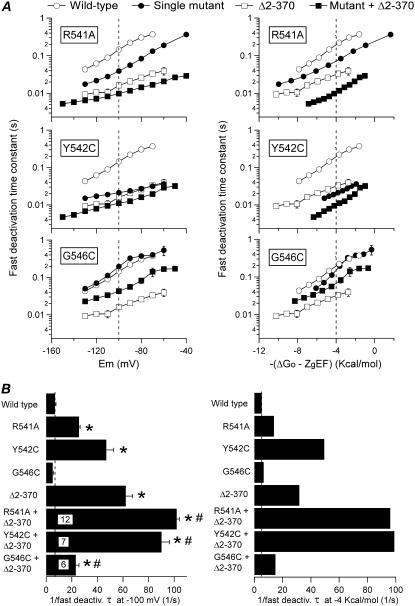
Gating kinetics and underlying thermodynamic properties of human ether-a-go-go-related gene (HERG) K(+) channels expressed in Xenopus oocytes were studied using protocols able to yield true steady-state kinetic parameters. Channel mutants lacking the initial 16 residues of the amino terminus before the conserved eag/PAS region showed significant positive shifts in activation voltage dependence associated with a reduction of z(g) values and a less negative DeltaG(o), indicating a deletion-induced displacement of the equilibrium toward the closed state. Conversely, a negative shift and an increased DeltaG(o), indicative of closed-state destabilization, were observed in channels lacking the amino-terminal proximal domain. Furthermore, accelerated activation and deactivation kinetics were observed in these constructs when differences in driving force were considered, suggesting that the presence of distal and proximal amino-terminal segments contributes in wild-type channels to specific chemical interactions that raise the energy barrier for activation. Steady-state characteristics of some single point mutants in the intracellular loop linking S4 and S5 helices revealed a striking parallelism between the effects of these mutations and those of the amino-terminal modifications. Our data indicate that in addition to the recognized influence of the initial amino-terminus region on HERG deactivation, this cytoplasmic region also affects activation behavior. The data also suggest that not only a slow movement of the voltage sensor itself but also delaying its functional coupling to the activation gate by some cytoplasmic structures possibly acting on the S4-S5 loop may contribute to the atypically slow gating of HERG.
Figures
References
-
- Sanguinetti, M. C., C. Jiang, M. E. Curran, and M. T. Keating. 1995. A mechanistic link between an inherited and an acquired cardiac arrhythmia: HERG channel encodes the IKr potassium channel. Cell. 81:299–307. - PubMed
-
- Trudeau, M. C., J. W. Warmke, B. Ganetzky, and G. A. Robertson. 1995. HERG, a human inward rectifier in the voltage-gated potassium channel family. Science. 269:92–95. - PubMed
-
- Barros, F., C. Villalobos, J. García-Sancho, D. del Camino, and P. de la Peña. 1994. The role of the inwardly rectifying K+ current in resting potential and thyrotropin-releasing hormone-induced changes in cell excitability of GH3 rat anterior pituitary cells. Pflugers Arch. 426:221–230. - PubMed
-
- Barros, F., D. del Camino, L. A. Pardo, T. Palomero, T. Giráldez, and P. de la Peña. 1997. Demonstration of an inwardly rectifying K+ current component modulated by thyrotropin-releasing hormone and caffeine in GH3 rat anterior pituitary cells. Pflugers Arch. 435:119–129. - PubMed
-
- Bauer, C. K., R. Schäfer, D. Schiemann, G. Reid, I. Hanganu, and J. R. Schwarz. 1999. A functional role of the erg-like inward-rectifying K+ current in prolactin secretion from rat lactotrophs. Mol. Cell. Endocrinol. 148:37–45. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
