

Cellular and molecular determinants targeting the Caenorhabditis elegans PHR protein RPM-1 to perisynaptic regions
- PMID: 18224716
- PMCID: PMC2657606
- DOI: 10.1002/dvdy.21446
Cellular and molecular determinants targeting the Caenorhabditis elegans PHR protein RPM-1 to perisynaptic regions
Abstract
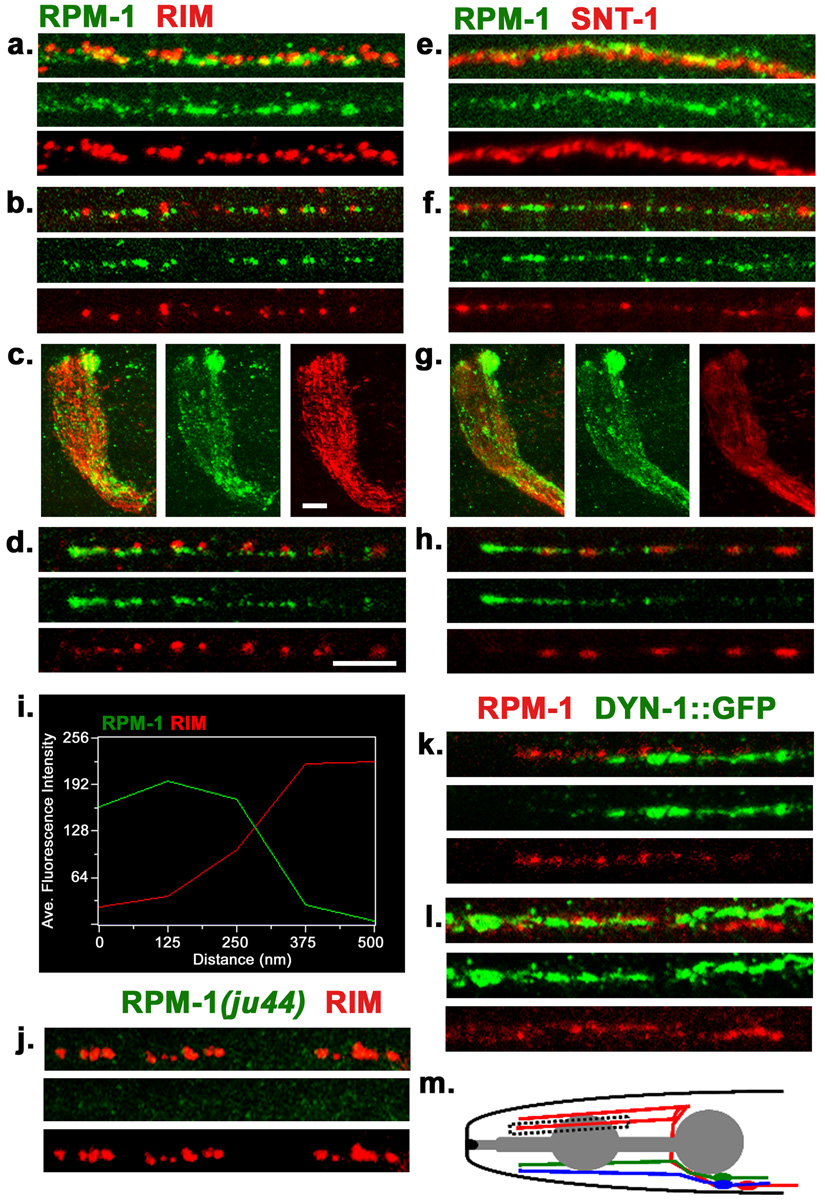
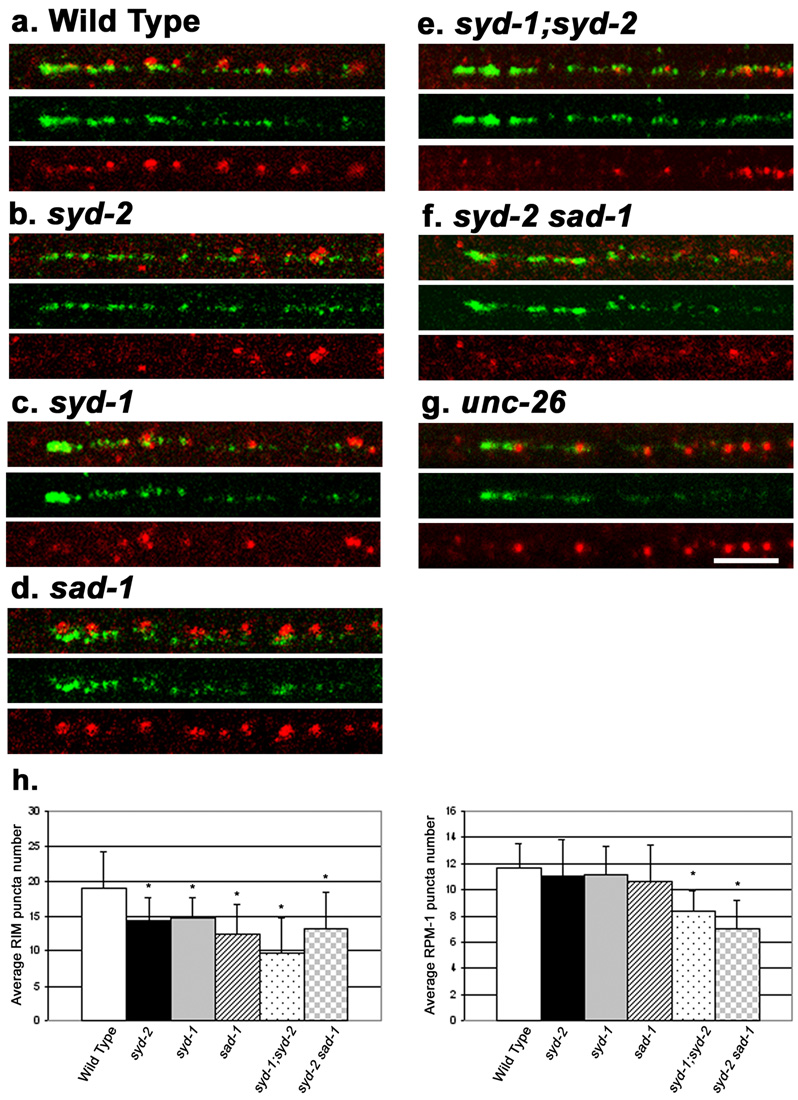
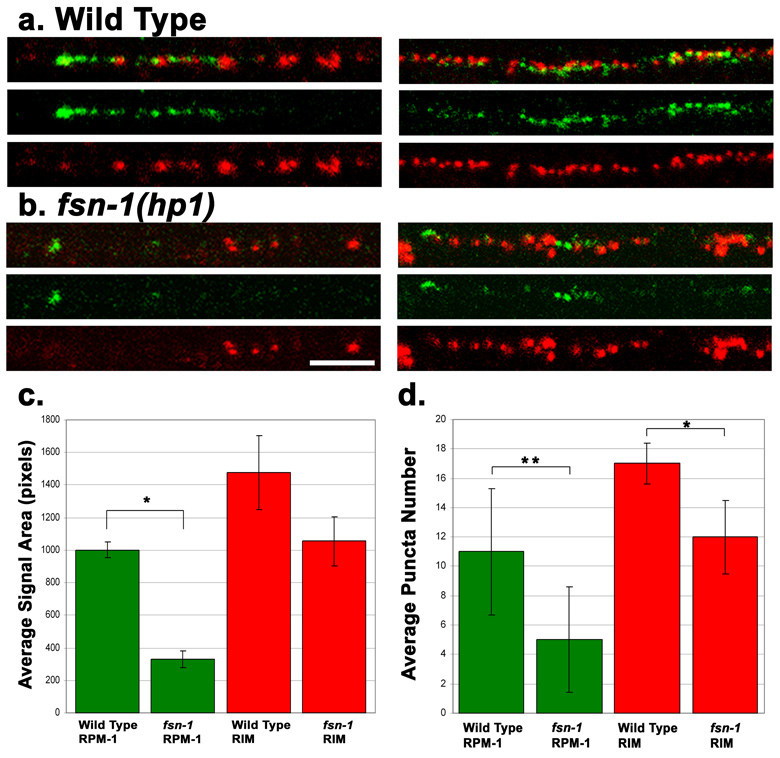
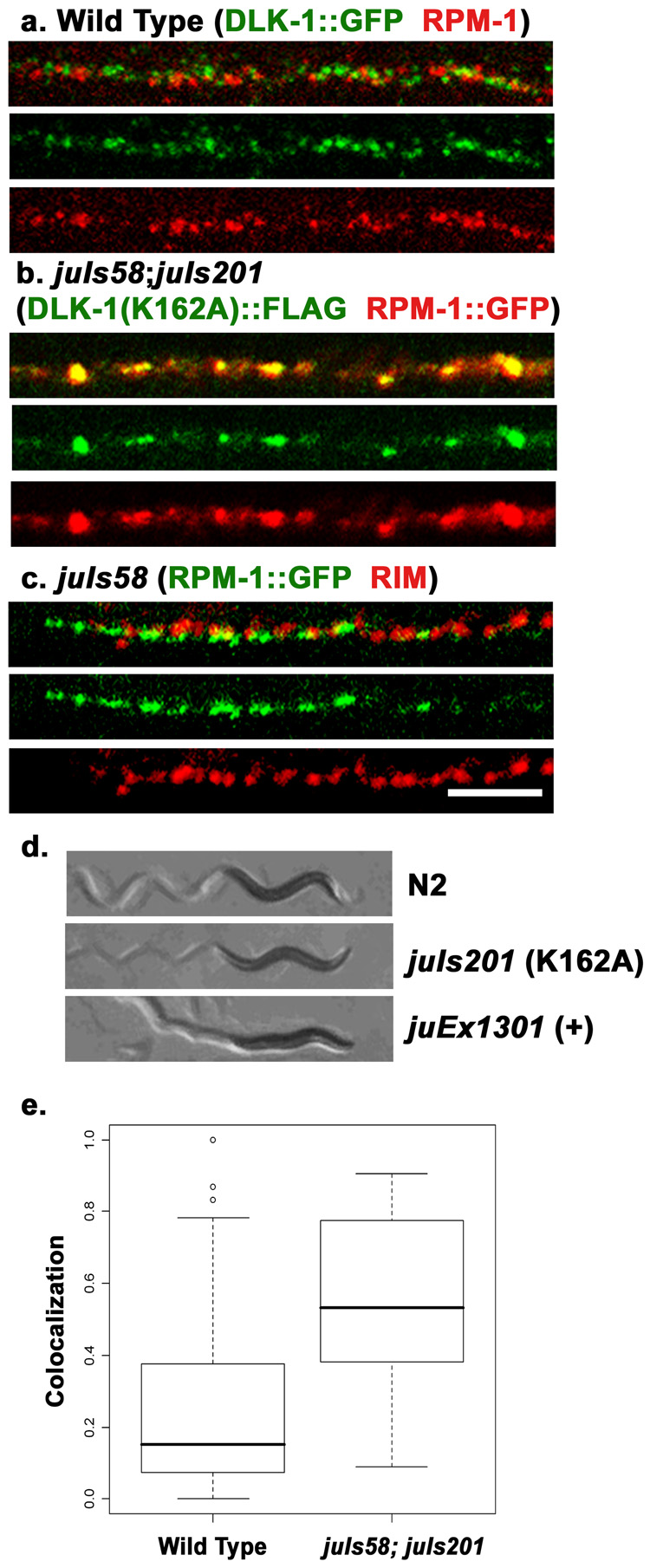
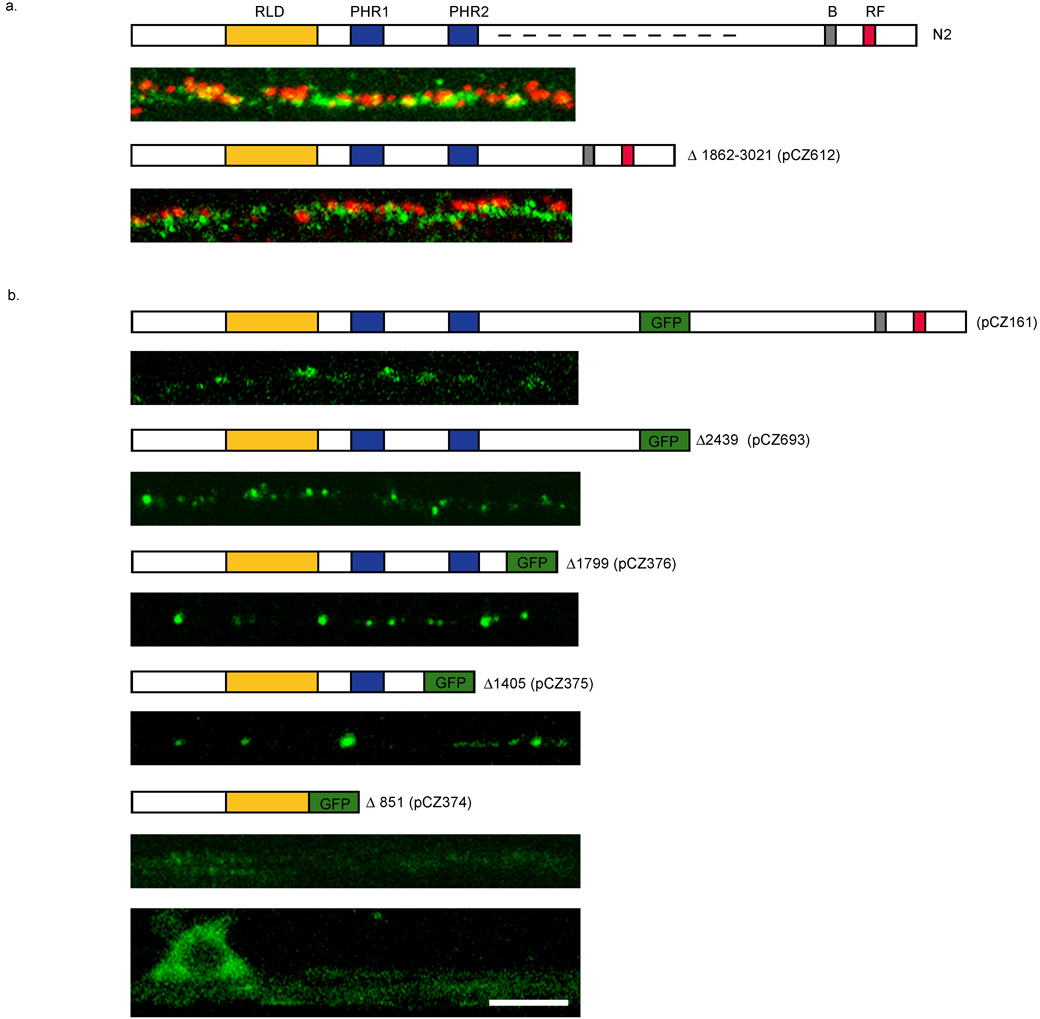
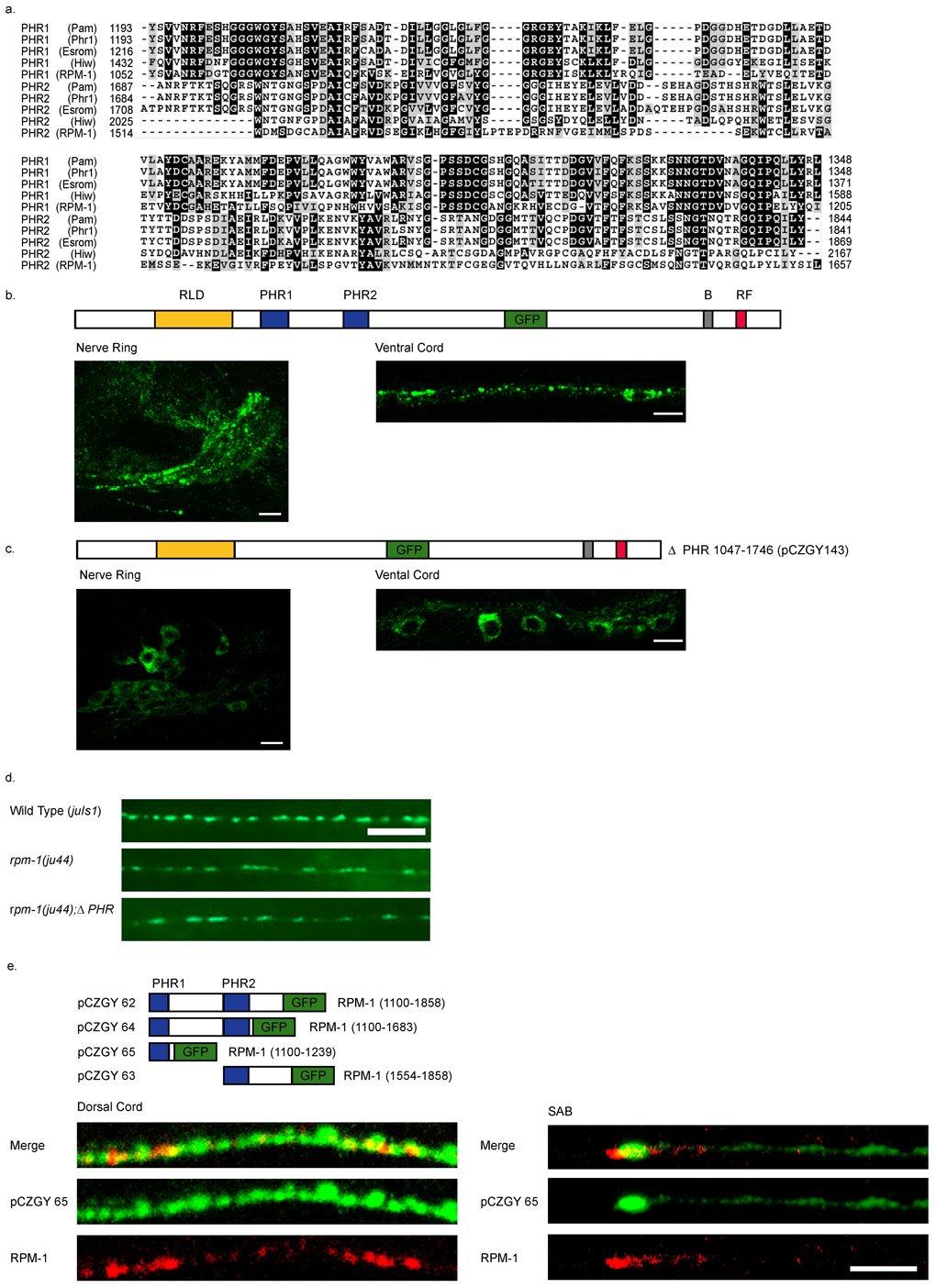
Caenorhabditis elegans RPM-1 is a member of a conserved protein family, the PHR proteins, that includes human Pam, mouse Phr1, zebrafish Esrom, and Drosophila Highwire. PHR proteins play important roles in the development of the nervous system. In particular, mutations in rpm-1 cause a disruption of synaptic architecture, affecting the distribution of synaptic vesicles and the number of presynaptic densities. Using antibodies against RPM-1, we determined the localization of the endogenous RPM-1 protein in wild-type and in several mutants that affect synaptic development. Our analyses show that, in mature neurons, RPM-1 resides in a distinct region that is close to, but does not overlap with, the synaptic exo- and endocytosis domains. The localization of RPM-1 occurs independently of several proteins that function in the transport or assembly of synapse components, and its abundance is partially dependent on its binding partner the F-box protein FSN-1. RPM-1 has been shown to target the MAPKKK DLK-1 for degradation. We show that activated DLK-1 may be preferentially targeted for degradation. Furthermore, using transgene analysis, we identified a critical role of the conserved PHR domain of RPM-1 in its subcellular localization.
(c) 2008 Wiley-Liss, Inc.
Figures
References
-
- Ackley BD, Harrington RJ, Hudson ML, Williams L, Kenyon CJ, Chisholm AD, Jin Y. The Two Isoforms of the Caenorhabditis elegans Leukocyte-Common Antigen Related Receptor Tyrosine Phosphatase PTP-3 Function Independently in Axon Guidance and Synapse Formation. J. Neurosci. 2005;25:7517–7528. - PMC - PubMed
-
- Coyle IP, Koh YH, Lee WC, Slind J, Fergestad T, Littleton JT, Ganetzky B. Nervous wreck, an SH3 adaptor protein that interacts with Wsp, regulates synaptic growth in Drosophila. Neuron. 2004;41:521–534. - PubMed
-
- Crump JG, Zhen M, Jin Y, Bargmann CI. The SAD-1 kinase regulates presynaptic vesicle clustering and axon termination. Neuron. 2001;29:115–129. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
