

Anandamide biosynthesis catalyzed by the phosphodiesterase GDE1 and detection of glycerophospho-N-acyl ethanolamine precursors in mouse brain
- PMID: 18227059
- PMCID: PMC2431036
- DOI: 10.1074/jbc.M707807200
Anandamide biosynthesis catalyzed by the phosphodiesterase GDE1 and detection of glycerophospho-N-acyl ethanolamine precursors in mouse brain
Abstract
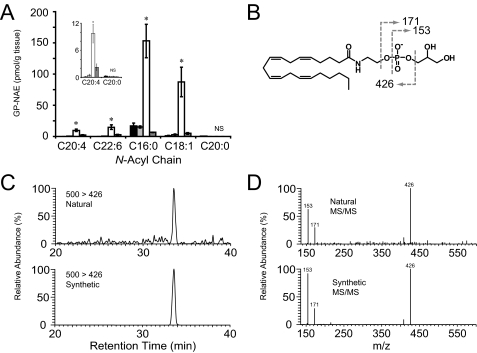
Anandamide (AEA) is an endogenous ligand of cannabinoid receptors and a well characterized mediator of many physiological processes including inflammation, pain, and appetite. The biosynthetic pathway(s) for anandamide and its N-acyl ethanolamine (NAE) congeners remain enigmatic. Previously, we proposed an enzymatic route for producing NAEs that involves the double-O-deacylation of N-acyl phosphatidylethanolamines (NAPEs) by alpha/beta-hydrolase 4 (ABDH4 or Abh4) to form glycerophospho (GP)-NAEs, followed by conversion of these intermediates to NAEs by an unidentified phosphodiesterase. Here, we report the detection and measurement of GP-NAEs, including the anandamide precursor glycerophospho-N-arachidonoylethanolamine (GP-NArE), as endogenous constituents of mouse brain tissue. Inhibition of the phosphodiesterase-mediated degradation of GP-NAEs ex vivo resulted in a striking accumulation of these lipids in brain extracts, suggesting a rapid endogenous flux through this pathway. Furthermore, we identify the glycerophosphodiesterase GDE1, also known as MIR16, as a broadly expressed membrane enzyme with robust GP-NAE phosphodiesterase activity. Together, these data provide evidence for a multistep pathway for the production of anandamide in the nervous system by the sequential actions of Abh4 and GDE1.
Figures







References
-
- Kuehl, F. A., Jacob, T. A., Ganley, O. H., Ormond, R. E., and Meisinger, M. A. P. (1957) J. Am. Chem. Soc. 79 5577-5578
-
- Bachur, N. R., Masek, K., Melmon, K. L., and Udenfriend, S. (1965) J. Biol. Chem. 240 1019-1024 - PubMed
-
- Natarajan, V., Reddy, P. V., Schmid, P. C., and Schmid, H. H. (1982) Biochim. Biophys. Acta 712 342-355 - PubMed
-
- Devane, W. A., Hanus, L., Breuer, A., Pertwee, R. G., Stevenson, L. A., Griffin, G., Gibson, D., Mandelbaum, A., Etinger, A., and Mechoulam, R. (1992) Science 258 1946-1949 - PubMed
-
- Cravatt, B. F., Giang, D. K., Mayfield, S. P., Boger, D. L., Lerner, R. A., and Gilula, N. B. (1996) Nature 384 83-87 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
