

Kinesin and dynein-dynactin at intersecting microtubules: motor density affects dynein function
- PMID: 18227130
- PMCID: PMC2275709
- DOI: 10.1529/biophysj.107.120014
Kinesin and dynein-dynactin at intersecting microtubules: motor density affects dynein function
Abstract
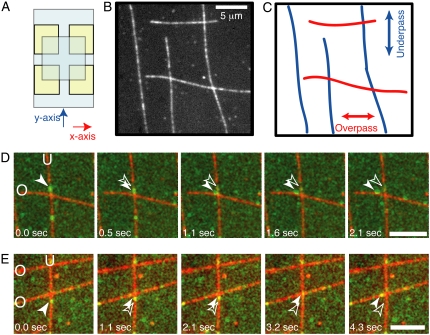
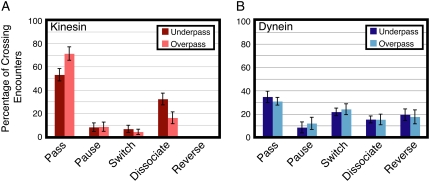
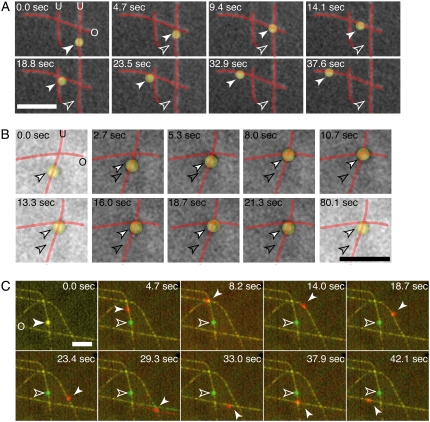
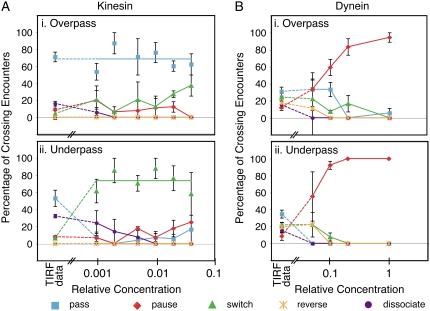
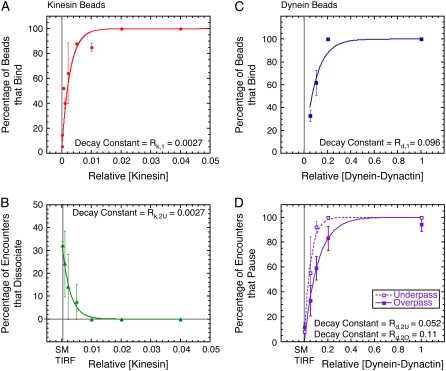
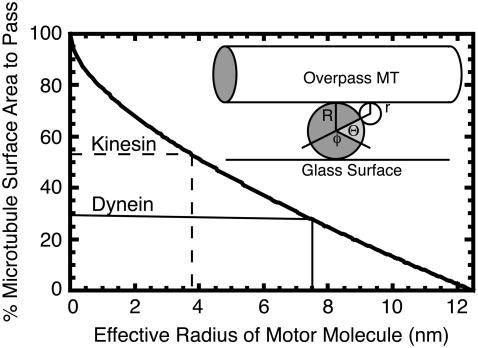
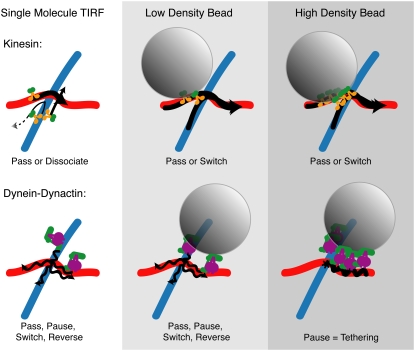
Kinesin and cytoplasmic dynein are microtubule-based motor proteins that actively transport material throughout the cell. Microtubules can intersect at a variety of angles both near the nucleus and at the cell periphery, and the behavior of molecular motors at these intersections has implications for long-range transport efficiency and accuracy. To test motor function at microtubule intersections, crossovers were arranged in vitro using flow to orient successive layers of filaments. Single kinesin and cytoplasmic dynein-dynactin molecules fused with green-fluorescent protein, and artificial bead cargos decorated with multiple motors, were observed while they encountered intersections. Single kinesins tend to cross intersecting microtubules, whereas single dynein-dynactins have a more varied response. For bead cargos, kinesin motion is independent of motor number. Dynein beads with high motor numbers pause, but their actions become more varied as the motor number decreases. These results suggest that regulating the number of active dynein molecules could change a motile cargo into one that is anchored at an intersection, consistent with dynein's proposed transport and tethering functions in the cell.
Figures
References
-
- Vale, R. D. 2003. The molecular motor toolbox for intracellular transport. Cell. 112:467–480. - PubMed
-
- Svoboda, K., C. F. Schmidt, B. J. Schnapp, and S. M. Block. 1993. Direct observation of kinesin stepping by optical trapping interferometry. Nature. 365:721–727. - PubMed
-
- Kon, T., M. Nishiura, R. Ohkura, Y. Y. Toyoshima, and K. Sutoh. 2004. Distinct functions of nucleotide-binding/hydrolysis sites in the four AAA modules of cytoplasmic dynein. Biochemistry. 43:11266–11274. - PubMed
-
- Burgess, S. A., M. L. Walker, H. Sakakibara, P. J. Knight, and K. Oiwa. 2003. Dynein structure and power stroke. Nature. 421:715–718. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
