

Variable gearing in pennate muscles
- PMID: 18230734
- PMCID: PMC2234215
- DOI: 10.1073/pnas.0709212105
Variable gearing in pennate muscles
Abstract
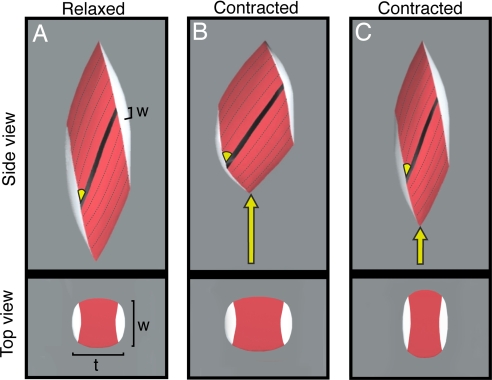
Muscle fiber architecture, i.e., the physical arrangement of fibers within a muscle, is an important determinant of a muscle's mechanical function. In pennate muscles, fibers are oriented at an angle to the muscle's line of action and rotate as they shorten, becoming more oblique such that the fraction of force directed along the muscle's line of action decreases throughout a contraction. Fiber rotation decreases a muscle's output force but increases output velocity by allowing the muscle to function at a higher gear ratio (muscle velocity/fiber velocity). The magnitude of fiber rotation, and therefore gear ratio, depends on how the muscle changes shape in the dimensions orthogonal to the muscle's line of action. Here, we show that gear ratio is not fixed for a given muscle but decreases significantly with the force of contraction (P < 0.0001). We find that dynamic muscle-shape changes promote fiber rotation at low forces and resist fiber rotation at high forces. As a result, gearing varies automatically with the load, to favor velocity output during low-load contractions and force output for contractions against high loads. Therefore, muscle-shape changes act as an automatic transmission system allowing a pennate muscle to shift from a high gear during rapid contractions to low gear during forceful contractions. These results suggest that variable gearing in pennate muscles provides a mechanism to modulate muscle performance during mechanically diverse functions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Biewener AA, Farley CT, Roberts TJ, Temaner M. J Appl Physiology. 2004;97:2266–2274. - PubMed
-
- Carrier DR, Heglund NC, Earls KD. Science. 1994;265:651–653. - PubMed
-
- Brainerd EL, Azizi E. J Exp Biol. 2005;208:3249–3261. - PubMed
-
- Gans C, Bock W. Ergeb Anat Entw Gesch. 1965;38:115. - PubMed
-
- Steno N. Elementorum Myologiae Specimen, seu Musculi Descripto Geometrica. Italy: Florence; 1667.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
