

Cooperative function of Tbx1 and Brn4 in the periotic mesenchyme is necessary for cochlea formation
- PMID: 18231833
- PMCID: PMC2536808
- DOI: 10.1007/s10162-008-0110-6
Cooperative function of Tbx1 and Brn4 in the periotic mesenchyme is necessary for cochlea formation
Abstract
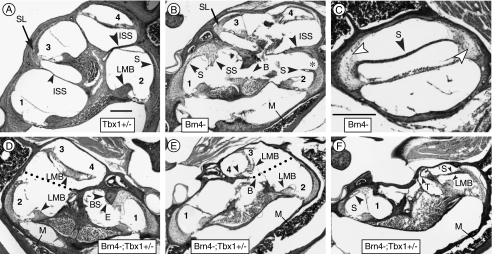
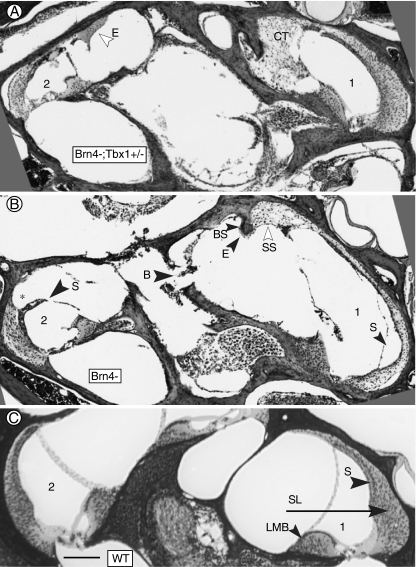
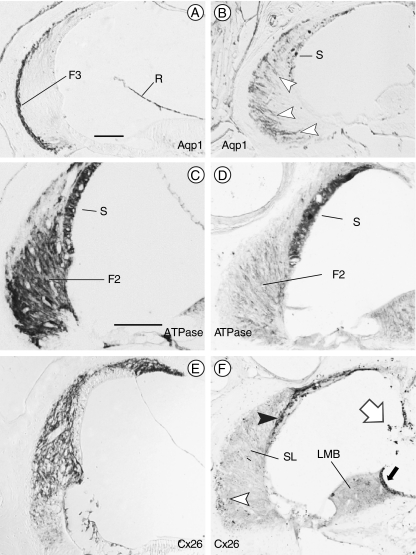
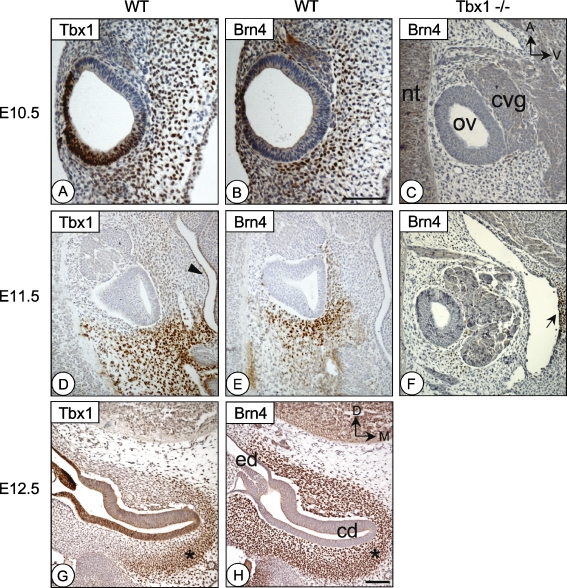
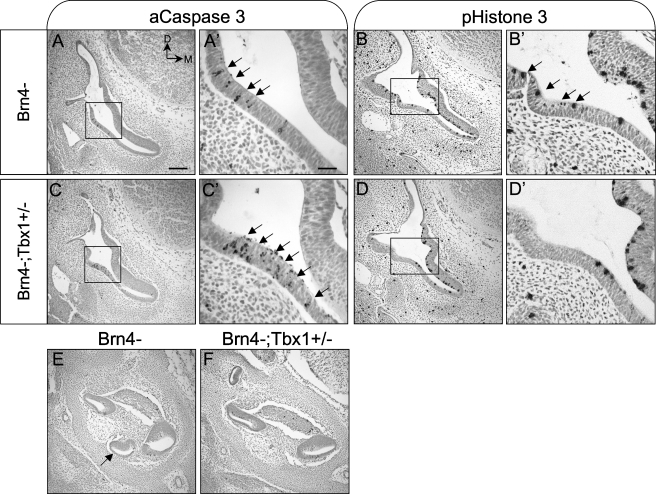
The T-box transcription factor TBX1 has been identified as the major gene responsible for the etiology of velocardiofacial syndrome/DiGeorge syndrome (VCFS/DGS). Conductive hearing loss occurs in a majority of patients with this syndrome, while sensorineural deafness has also been reported in some cases. Mutations in POU3F4/BRN4, a POU domain transcription factor, cause DFN3, an X-linked nonsyndromic form of deafness characterized by mixed conductive and sensorineural hearing loss. Inactivation of the murine orthologues of these genes causes similar defects to those seen in humans and has provided excellent models for the study of inner ear development. Tbx1 and Brn4 are expressed in the mesenchymal cells surrounding the otic vesicle and have been shown to play roles in cochlear outgrowth. Furthermore, expression of Brn4 is reduced in Tbx1 null mutants, suggesting a possible genetic interaction between these genes. To test whether Tbx1 and Brn4 function in a common pathway, mice mutant for both genes were generated and analyzed for inner ear defects. Brn4-;Tbx1+/- mutants displayed a significant reduction in the number of turns of the cochlea compared to Brn4- or Tbx1+/- mice. In addition, Brn4-;Tbx1+/- mice displayed structural defects in the apical cochlea indicative of Mondini dysplasia found in patients with either VCFS/DGS or DFN3. These data establish a genetic interaction between Tbx1 and Brn4 relevant to human disease and indicate a function of these genes in signaling from the periotic mesenchyme to the otic vesicle to direct proper coiling of the cochlear duct.
Figures
References
-
- Arellano B, Ramirez Camacho R, Garcia Berrocal JR, Villamar M, del Castillo I, Moreno F. Sensorineural hearing loss and Mondini dysplasia caused by a deletion at locus DFN3. Arch. Otolaryngol. Head Neck Surg. 126:1065–1069, 2000. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
