

A dormant internal ribosome entry site controls translation of feline immunodeficiency virus
- PMID: 18234788
- PMCID: PMC2268504
- DOI: 10.1128/JVI.02038-07
A dormant internal ribosome entry site controls translation of feline immunodeficiency virus
Abstract
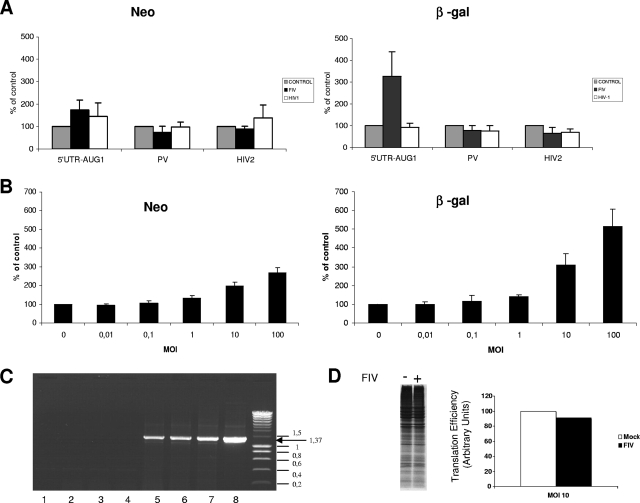
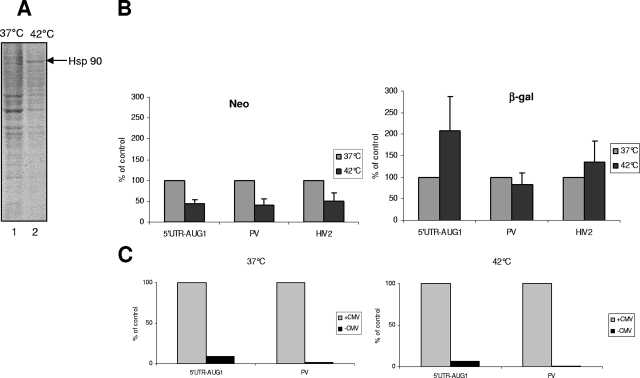
The characterization of internal ribosome entry sites (IRESs) in virtually all lentiviruses prompted us to investigate the mechanism used by the feline immunodeficiency virus (FIV) to produce viral proteins. Various in vitro translation assays with mono- and bicistronic constructs revealed that translation of the FIV genomic RNA occurred both by a cap-dependent mechanism and by weak internal entry of the ribosomes. This weak IRES activity was confirmed in feline cells expressing bicistronic RNAs containing the FIV 5' untranslated region (UTR). Surprisingly, infection of feline cells with FIV, but not human immunodeficiency virus type 1, resulted in a great increase in FIV translation. Moreover, a change in the cellular physiological condition provoked by heat stress resulted in the specific stimulation of expression driven by the FIV 5' UTR while cap-dependent initiation was severely repressed. These results reveal the presence of a "dormant" IRES that becomes activated by viral infection and cellular stress.
Figures





References
-
- Belsham, G. J., and N. Sonenberg. 2000. Picornavirus RNA translation: roles for cellular proteins. Trends Microbiol. 8330-335. - PubMed
-
- Brasey, A., M. Lopez-Lastra, T. Ohlmann, N. Beerens, B. Berkhout, J.-L. Darlix, and N. Sonenberg. 2003. The leader of human immunodeficiency virus type 1 genomic RNA harbors an internal ribosome entry segment that is active during the G2/M phase of the cell cycle. J. Virol. 773939-3949. - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
